Ramenné pásy cicavec zjednodušený pozostáva z veľkého lopatkyktoré majú rast kakaových kyselín a kľúčna kosť(obr. 80, 81). V embryonálnom štádiu vývoja au dospelých monotremes zvieratá coracoid je nezávislá kosť.
ramenoje umiestnený na hrudi, má trojuholníkový tvar a vysoký vzrast (hrebeň) akro- miónproces pripevnenia svalov. V mieste kokosového procesu a akromionového procesu lopatky je kĺbová fossa pre pripojenie k prednej končatine.
kľúčna kosťtyčový tvar slúži na spojenie ramena a hrudnej kosti. Klíčnica je najviac vyvinutá u zvierat, ktoré svojimi prednými končatinami vykonávajú zložité pohyby (netopiere, krtky, primáty, hlodavce atď.). V mnohých druhoch, ktoré sa vyznačujú rovnomerným pohybom (kopytníky, psi), nie je kľúčná kosť. Všeobecne je pletenec predných končatín spojený so svorkami a väzmi prostredníctvom axiálnej kostry.
Panvový opasok (Obr. 82 A) pozostáva z párov bezmenné kostitvorené kĺbovým spojením ilea, sedacieho a ochlpeniakostí. Panva uzavretého typu a cez iliakálne kosti je pevne spojená sakrálne oddelenie chrbtice. Keď prvky panvy rastú spolu verejná fúzia a formované acetabulum na spojenie zadných končatín. V dolnej časti kosti bez názvu je pridržiavací otvor.
Kostra párových končatín
Konce cicavcov, rovnako ako končeky ostatných suchozemských stavovcov, sú päťbodového typu a predstavujú páku s tromi koncami, z ktorých všetky časti sú pohyblivo prepojené.
predné končatiny pozostáva z rameno, predlaktie a ruka (Obr. 81). ramennej dobre vyvinutý, má zaoblenú hlavu na pripojenie k opasku končatiny. predlaktia tvoria dve kosti - radiálne a ulnárnemajúce olecranon hore. Na križovatke ramena a predlaktia je lakťový kĺb.
štetec zahŕňa tri oddelenia - zápästie, metakarpus a falanga. zápästia pozostáva z 8–10 prvkov usporiadaných do troch radov , Metacarpus obsahuje päť kostí (podľa počtu prstov). Každý prst má tri falangy, s výnimkou prvého prsta, ktorý sa skladá z dvoch falong. Na rozdiel od interkarpálneho členenia plazov a vtákov, mobil kĺb nachádza sa u cicavcov medzi predlaktím a rukou (ako obojživelníci).
Zadná končatina cicavce majú tiež tri divízie - stehná, holene a chodidla(Obr. 82 B). Femur kosť masívna a zasunutá okrúhla hlava do acetabula panvového pletenca. Palička obsahuje dve kosti - veľká holenná kosť a malá holenná kosť.Väčšia tibiálna kosť predstavuje vnútorný prvok holennej kosti a fibula je jej vonkajšou časťou.
Medzi stehnom a dolnou končatinou je kĺbzakryté jabĺčkoktorý je evolučne formovaný ich osifikovanými svalovými šliach. Povrchy kĺbových kostí sú pokryté chrupavkovými útvarmi. Členkový kĺb cicavce umiestnené medzi dolnými koncami holennej kosti holennej kosti a proximálnou nohou.

Obr. 82. Panvový opasok (A) a zadné končatiny (B) líšky:
1 – iliakálna kosť, 2 - ischiálna kosť, 3 - ochlpenie, 4 - acetabulum, 5 - stehenná kosť, 6 - patella, 7 - holenná kosť, 8 - holenná kosť, 9 - tarzus, 10 - tarzus, 11 - falangy prstov , 12 - členková kosť, 13 - kosť päty
Tarzálne prvky chodidlá sú usporiadané do troch radov a tvoria vyrastanie - pätnej kosticharakteristika cicavcov. priehlavok pozostáva z piatich kostí, ku ktorým sú pripevnené falangy prstov. Rovnako ako v predných končatinách sú na každom prsteň tri, s výnimkou prvého (vnútorného), ktorý má iba dve falangy.
Cicavce sa vyznačujú špeciálnym nastavením končatín vzhľadom na telo: sú umiestnené pod telom, čo prispieva k rozvoju väčšej ovládateľnosti pohybov. V tomto prípade je lakťový kĺb nasmerovaný dozadu a kolenný kĺb, naopak, je orientovaný predne.
Kvôli druhovej diverzite cicavcov, ich životnému štýlu a spôsobu pohybu, relatívnej dĺžke končatín, pomeru základných prvkov, konfigurácii kostí, počtu prstov atď., Sa napríklad netopiere prudko zvyšujú v dĺžke prednej končatiny, najmä zamestnancov na pripevnenie lietajúcej membrány.
Naopak, predná časť krtka je skrátená, dlaňová časť ruky je roztiahnutá a šiesty prst je dobre vyvinutý. U veľryb sa kefa prudko predlžuje v dôsledku zvýšenia počtu falangov na prstoch, v dôsledku čoho sa končatina získala veľmi podobná plutve plutvy. Výrazne zmenšené a pozmenené prsty zvierat s veľkou biomasou a rýchlym chodom (vzpriamene, kopytnaté).
Chrbtica pozostáva z krčných, hrudných, bedrových, sakrálnych a kaudálnych oblastí. Jeho charakteristickým znakom je tvar platielis (s rovnými povrchmi) stavcov, medzi ktorými sú umiestnené chrupavkové medzistavcové platničky. Dobre definovaný horný oblúk. V krčnej oblasti je sedem stavcov, ktorých dĺžka závisí od dĺžky krku; iba mravec a lenivosť - Choloepus hof-fmani majú 6 z nich a lenivosť - Bradypus 8-10. Krčné stavce sú veľmi dlhé u žirafy a veľmi krátke u veľrýb bez krčka. K stavcom hrudnej oblasti sú pripevnené hrany, ktoré sa tvoria hrudná klietka, Jeho hrudná kosť je plochá a má len u netopierov a druhov nory, ktoré majú silné predné končatiny (napríklad krtky), má malý hrebeň (kýl), ktorý slúži ako miesto pripojenia hrudných svalov. V oblasti hrudníka 9–24 (zvyčajne 12–15) stavcov nesie posledné 2–5 hrudných stavcov pozitívne rebrá, ktoré nedosahujú hrudnú kosť. Základné rebrá sa spájajú s ich veľkými priečnymi procesmi. Sakrálnu oblasť tvoria 4-10 zarastené stavce, z ktorých iba prvé dve sú skutočnými sakrálnymi a zvyšok sú chvosty. Počet voľných kaudálnych stavcov sa pohybuje v rozmedzí od 3 (v gibbone) do 49 v jašterici dlhoocasej.
Stupeň mobility jednotlivých stavcov je odlišný. U malých bežiacich a lezúcich zvierat je veľká, po celej dĺžke chrbtice, takže sa jej telo môže ohýbať rôznymi smermi a dokonca sa skrúti do gule. Hrudné a bedrové stavce sú menej mobilné u veľkých, rýchlo sa pohybujúcich zvierat. U cicavcov, ktoré sa pohybujú po zadných nohách (klokani, jerboi, mikiny), sa najväčšie stavce nachádzajú pri spodnej časti chvosta a krížovej kosti a ich hodnota vpred sa postupne znižuje. Naopak, u kopytníkov sú stavce a najmä ich spinálne procesy väčšie pred hrudnou oblasťou, kde sú k nim pripevnené silné svaly krku a čiastočne predné končatiny (
Jednou z čŕt kostry cicavcov je veľký vývoj chrupavky u mladých zvierat a jej zachovanie počas postembryonického obdobia vývoja. Je to kvôli zvláštnostiam rastu chrupavky a kostí v dôsledku rastu samotného zvieraťa. Je to chrupavka, ktorá rastie s celou jej hmotou (intersticiálne), rast nastáva rýchlo a proporcie častí sa môžu udržiavať, zatiaľ čo kosť rastie pomalšie a iba z povrchu, takže časti kostry, ktoré sa objavujú, vždy zostávajú v rovnakej vzdialenosti a pomer častí rastie. nemožné. Preto mladé zvieratá vždy obsahujú veľa chrupaviek, najmä v miestach, kde je silný rast. Dlhé kosti končatín sa teda vždy skladajú zo strednej časti - diafýzy, ktorá predo všetkým osifikuje, a koncov, ktoré chrupavka dlho chráni - epifýzy. Osifikácia diafýzy sa začína predovšetkým periférnym perichondriálnym, to znamená na úkor buniek pochádzajúcich zo spojivového tkaniva, ktoré opotrebováva chrupavku, z perichondrium; potom sa k nemu pripojí vnútorná osifikácia chrupavky - endochondrálna osifikácia; až neskôr, keď sa diafýza objaví dezintegráciou dutiny kostnej drene, epifýzy začínajú osifikovať endochondriálne; medzi diafýzou a epifýzou však chrupavka zostáva dlho, s pomocou ktorej sa uskutočňuje rast kostí, pričom sa zachovávajú proporcie (obr. 530).

chrbtica cicavce, na rozdiel od Sauropsidy, sa vyznačujú neprítomnosťou voľných kĺbov medzi stavcami, s výnimkou prvého a druhého stavca. Medzistavcové elastické spojivové tkanivové disky ležia medzi stavcami. Spravidla to vedie k menšej flexibilite chrbtice, čo je dôsledkom špecializovanejšieho pohybu cicavcov s končatinami.
K osifikácii stavcov dochádza prostredníctvom epifýzy chrupavky. Tela má buď rovné alebo slabo vypuklé povrchy, vo väčšine moderných kopytníkov sú mierne opistocelous. Obratle sú dobre vyvinuté, najmä v oblasti hrudníka, v spinálnych procesoch, ako aj v priečnych procesoch, ktoré sa nachádzajú na oblúkoch; na jeho základni sú kĺbové procesy.
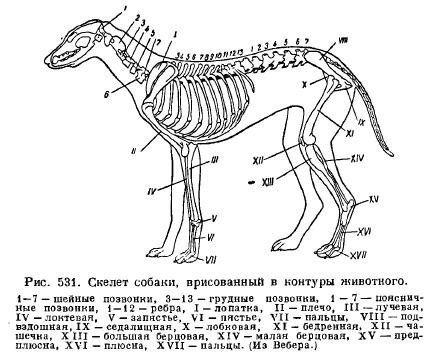
Počet krčných stavcov je zvyčajne 7 (obr. 531), bez ohľadu na dĺžku krku, takže dĺžka krku, na rozdiel od vtákov, závisí výlučne od dĺžky stavcov. Iba krkavec (Manatus) a jeden druh lenivosti (Choloepus hoffmani) majú 6 krčných stavcov, zatiaľ čo trojprstá lenivosť (Bradypus) má 9 z nich. zatiaľ čo v krku žirafy je tých istých 7 stavcov veľmi dlhých. Prvé 2 krčné stavce sú upravené na atlas a epistrofiu, spojené skutočnými kĺbmi a pri jednorazovom dentálnom procese stále fungujú ako špeciálna kosť a nerastú spolu s telom epistrofie ako u iných cicavcov. Základné rebrá krčnej oblasti sa fúzujú so stavcami, v dôsledku čoho sa tvoria perforované priečne procesy. U veľrýb sa vyskytuje čiastočné alebo úplné uzdravenie krčných stavcov; v jerbooch rastú všetky krčné stavce spolu s výnimkou siedmich, v marsupiálnom móle (Notoryctes) zlúčenom od 2. do 6., v mole, od 3. do 4..
Počet hrudných stavcov, ktoré sa vyznačujú prichytením rebier k nim, sa pohybuje od 9 [v jednom veľrybe (Hyperoodon) a v ramene (Tatusia)] do 25 [v lenivosti], zvyčajne je to 12–13. Procesy hrudných stavcov sú dobre vyvinuté: spinálne (proc. Spinosus), priečne alebo diapophysis (proc. Transversus), artikulárne alebo zygapofyzy (proc. Articularis); rebrá sú kĺbovo spojené hlavami k telu jedného alebo dvoch susedných stavcov a tuberkúl k priečnym procesom. Skladajú sa z dvoch častí: kostného rebra - proximálneho a chrupavkového - distálneho. K hrudnej kosti sa dostanú iba rebrá predných hrudných stavcov, zadné rebrá končia voľne (nepravé rebrá). Bedrové stavce, ktoré sa vyznačujú neprítomnosťou rebier, ktoré sú obsiahnuté v zložení priečnych procesov u cicavcov od 2 do 9, sú obvykle 6-7.
Sakrálne stavce cicavcov sú tie stavce, ktoré sa spoja a kĺbovo spoja iliakálna kosť, Takéto stavce sú 1 až 10, ale iba 1 až 2 z nich sú skutočné sakrálne stavce, to znamená tie, v ktorých kĺbové spojenie s panvou nastáva nezávislou osifikáciou zodpovedajúcou sakrálnym rebrám; zvyšok sakrálnych stavcov je nepravdivý, to znamená, kaudálne stavce pripojené k krížovej kosti.
Počet stavcov vstupujúcich do chvostovej sekcie sa veľmi líši: od 3 (netopiere, gibbon) do 49 [jašterica (Manis macrura)].
hrudná kosť (sternum) cicavcov sa vyvíja fúziou brušných koncov rebier, ktoré potom tvoria jednu nepárovú doštičku pozostávajúcu z niekoľkých divízií (obr. 532).

V spojení s kľúčnou kosťou (ak existuje) as prvým párom rebier sa predĺžená predná časť hrudnej kosti nazýva grip (praesternum, manubrium); Nasleduje časť nazývaná telo (mezosternum, korpus), ktorá sa často rozkladá na sériu kostí, ktoré sa postupne nachádzajú medzi po sebe idúcimi pármi rebier; ďalej späť leží tzv. xiphoidný proces (xiphisternum). Zďaleka nie sú všetky rebrá spojené s hrudnou kosťou, viac alebo menej významný počet falošných rebier končí voľne (obr. 533).

U kloakálnych cicavcov (Monotremata) sa v dôsledku prítomnosti coracoids, ktoré nie sú prítomné u všetkých ostatných cicavcov, chrupavka predhrudia (prosternum) položí pred hrudnú žľazu, ktorá sa potom nahradí vyvíjajúcou sa kosťou, epigramom (episternmn), a chorcoidy, ktoré sa nahradia chorcoidmi, sa kostra kostra vymení. plazy.
Lebka cicavce majú množstvo rysov odlišných od plazov lebky. Tieto rysy sa však neobjavili okamžite, ale pomocou postupnej zmeny, s ktorou sa dá človek oboznámiť štúdiom lebiek fosílnych živočíšnych tvorov (Theromorpha), najmä z poriadku zvieracích zubov (Theriodontia). Cicavčia lebka je lebka tropibazálna, ale so skrátenou interorbitálnou časťou, vďaka ktorej vyzerá ako platybasal, s veľkou lebkou, s výraznou tendenciou rásť jednotlivé kosti spolu,
Umiestnenie kostí v lebke cicavcov sa môže posudzovať podľa priloženého diagramu (Obr. 534).
Na laterálnych týlnych kostiach, zvonka od kĺbových hľúz, sa obvykle vyskytuje týlny proces (proc. Paroccipitalis), zvlášť dobre vyvinutý u hlodavcov a u mnohých kopytníkov. Pred artikulárne tuberkulózy je obvykle otvor na výstup z hypoglossálneho (XII) nervu (pre. Condyloideum anterius).
V chrupavke sluchovej kapsuly sa kladie niekoľko osifikácií, ktoré sa však veľmi skoro zlúčia do dvoch kostí: v prednej časti ležia - kamenisté (petrosum) a ležiace zvonku a zozadu - mastoidy (mastoideum); obe tieto kosti, ktoré spolu rastú, dávajú kamenistý mastoid alebo jednoducho kamennú kosť (petromastoideum, petrosum alebo perioticum), ktorá uzatvára vnútorný zvukový labyrint. Periotikum je ohraničené v zadnej časti hlavnými a bočnými týlnymi kosťami av prednej časti sféenoidmi a krídlami sfenoidné kosti; tak medzi týmito kosťami a kamenná kosť cievy a nervy prechádzajú, potom zostávajú výrazné medzery: za ňou je zadná roztrhnutá diera (pre. lacerum posterius) alebo jugulárna diera (pre. jugulare) pre priechod glossopharyngálneho (IX) nervu, vetvy nervu vagus (X) a pre príslušenstvo (XI) ) nervy, ako aj pre vnútornú krčnú žilu a pred ňou predný roztrhaný otvor (pre. lacerum anterius), ktorým prechádza vnútorná krčná tepna do lebky. Niekedy komunikácia skalnaté kosti s lebkou je taká slabá, že ju možno ľahko oddeliť spolu s ňou pripevnenou tympanickou kosťou alebo dokonca aj vtedy, keď macerácia lebky úplne odpadne, ako je to v prípade veľrybotvarých cicavcov.
Mimo ušnej oblasti sa tvoria dve ďalšie kosti. Jeden z nich, ktorého pôvod ešte nie je dostatočne jasný, sa vyvíja v oblasti vonkajšieho zvukovodu vo forme polkruhovej malej kosti. Je to tympanická kosť (tympanikum), ktorá je pravdepodobne homológom uhlovej kosti (angulare) čelenky plazov, ktorá prešla do príušnej oblasti spolu s ostatnými kosťami čelenky v dôsledku zmien v čeľustnom kĺbe. Keďže je spočiatku umiestnená vo forme polkruhovej alebo prstencovej kosti v oblasti ušného kanálika, často rastie a tvorí sa, zvyčajne s kamennou kosťou a niekedy so šupinatým vonkajším zvukovým kanálikom (meatus auditorius externus). Tympanická membrána je natiahnutá na tympanickej kosti. Hlbšie dovnútra sa tympanická kosť môže tiež podieľať na tvorbe stien stredného ucha. Tympanická kosť na spodnej strane sluchovej oblasti často napučí do tenkostenného mechúra (bulla ossea).
Druhou kosťou kožného pôvodu, ktorá sa podieľa na tvorbe kostry ušnej oblasti, je šupinatá kosť (skvamózna); u cicavcov získal výnimočný vývoj a význam v súvislosti s pripevnením dolnej čeľuste na ňu. U opíc a ľudí sa všetky tieto kosti ušnej oblasti navzájom spájajú a vytvárajú komplexnú časovú kosť (temporale); šupinatá kosť je tu iba časť, konkrétne squama temporalis časná kosťzatiaľ čo tympanická kosť je tympanická časť (pars tympanicum) a skutočné príušné chrupavkové kosti tvoria časti kamenisté (pars petrosa) a mastoidy (pars mastoidea). Stupeň vývoja šupinatej kosti a jej účasť na tvorbe stien mozgovej schránky a sluchového svalu sú veľmi rozdielne, ale šupinatá kosť má vždy kĺbový povrch (cavitas glenoidea) na spojenie s čelenkou. U cicavcov je teda na rozdiel od všetkých ostatných stavovcov veľmi originálne uchytenie dolnej čeľuste: nie k štvorcovej kosti, ale k šupinatej kosti lebky. Kĺbová plocha dolnej čeľuste leží na základe zygomatického procesu (proc. Zygomaticus) vychádzajúceho zo stupnice smerom von a dopredu. Pri jednorazovom prechode cez kĺbový povrch preniká dočasný kanál (can. temporalis), zvlášť dobre vyjadrený v platypus. Za kĺbovou fossou je proces (proc. Postglenoideus).
Pred sluchovým oddelením tvorí hlavicu lebka sfenoidná kosť (basicphenoidale), na vnútornom povrchu ktorého je viac alebo menej výrazná depresia - tzv. turecké sedlo (sella turtica), v ktorom je umiestnená hypofýza. Na stranách hlavnej klinovitej kosti ležia dolné časti prednej steny lebky, krídlové sfenoidné kosti (alisphenoidea), ktoré sa s ňou zvyčajne spájajú do jednej kosti, čím sa vytvárajú veľké krídla (alae magnae) tejto kosti. Kostné krídla vybavené dvoma otvormi: ovál (pre ovál) pre priechod tretej vetvy trigeminálneho nervu (V3) a guľatý (pre rotundum) pre priechod druhej vetvy toho istého nervu (V2). Prvý sa však často spája s prednou členitou dierou (pre. Lacerum anteris) a druhý sa často spája s klinovým orbitálnym otvorom (pre. Sphenorbitale), ktorý je súčasne obmedzený ako krídlová sféenoidná kosťa ležiace pred jej orbitálnou sfenoidnou kosťou. Očné nervy (okulomotorius III), blokáda (trochlearis IV), abducény VI a prvá vetva trigeminálneho nervu prechádzajú touto dierou.
Kraniálne veko je pokryté parietálnymi kosťami (parietale), ktoré často spolu rastú (v monotrónoch, niektorých vačkových a kopytníkov); medzi nimi a hornými týlnymi kosťami sa položia ďalšie dve kosti, ktoré sa zlúčia do jednej medzikostnej kosti (medzipriestorové), ktorá sa v niektorých oddeleniach (veľryby, vačkovce, damany, aardvark, vlna krídla a niektoré hlodavce) zachováva ako samostatná kosť. alebo s parietálnymi (u niektorých hlodavcov, sirén a prežúvavcov) alebo s hornou týlnou kosťou (u mäsožravcov, monopedov, primátov).
Pred hlavnou sfénoidnou kosťou leží predná sfenoidná kosť (praesphenoideum) s priehlbinou pre priehlbinu orbitálnych nervov; Na jej stranách, ktoré spolu s ňou zvyčajne rastú, ležia orbitálne klinovité kosti (orbitosphenoidea) s dierou pre priechod orbitálnych nervov (pre. Opticum), nazývané v prípade narastania prednými a hlavnými klinmi v tvare malých krídiel (alae parvae). Ako je uvedené vyššie, medzi oboma pterygoidmi na každej strane sú štrbinové otvory, cez ktoré prechádzajú nervy očných svalov a prvá vetva trigeminálneho nervu (pre. Sphenorbitale).
Na vrchole tejto rady kostí sú krycie predné kosti (predné) - zvyčajne dobre vyvinuté spárované kosti, niekedy vzájomne zarastené a nesúce sa v stonkách hovädzieho dobytka a na jeleňových konopných rohoch. Krycie lebky, predné kosti sú obmedzené nad a na vnútornej strane obežnej dráhy, často vytvárajú procesy: preorbitálne (proc. Praeorbitalis a za orbitálne (proc. Postorbitalis)) Niekedy okolo očí sa tvoria v dôsledku spojenia zygomatickej kosti (jugale) s frontálnou kosťou okolo orbitálneho kruhu ( niektoré hmyzožravce a mäsožravce, prežúvavce, monopody, napoly opice.) U opíc proces separácie očnej dutiny od dočasnej prebieha tak ďaleko, že medzi očnou dutinou a dočasnou dutinou sa vytvára kostná stena.
V prednej stene obežnej dráhy je malá inkrementálna slzná kosť (lacrymale), s dierou v slznom kanáli (pre. Lacrymale).
V nosovej oblasti je v chrupavkovej nosovej kapsule uložených niekoľko osifikácií. Najprv sa pozdĺž chrupavkového nosného septa v strede vyvinie kostné septo (lamina perpendicularis), ktoré zodpovedá strednej čuchovej kosti (mesethmoideum); po druhé, v zadnej stene nosnej kapsuly a v prednej stene mozgovej dutiny sa vyvíjajú dve bočné platne, ktoré sa prepichujú početnými [iba v kačici], tieto otvory sa nahradia jedným (pre. olfactorium)] otvormi pre priechod čuchového nervu - mrežovými platničkami (lamina cribrosa). bočné etmoidálne kosti (ethmoidalia lateralia). Z nosového septa prechádza osifikácia od bočných strán po stranách, vedie pozdĺž bočných stien nosných kapsúl a končí na spodnej strane nosa a vytvára početné výčnelky zo strán vo vnútri nosovej dutiny. Sú to čuchové škrupiny (ethmoturbinalia), ktoré spolu s bočnými doskami a dutinami bunkovej štruktúry niekedy tvoria veľmi komplexný nosný labyrint. Poloha dutín je zrejmá z priloženého schematického výkresu (Obr. 535). Z nosnej kosti (nosovej kosti), pokrývajúcej túto oblasť zhora, viste dolu do nosovej dutiny, tiež po jednej na každej strane, škrupiny (nazoturbmalia). Spodok nazálneho septa sa prekrýva s nespárovanou kosťou, ktorá sa predtým považovala za vomér (vomér). Nedávne štúdie však ukázali, že okrem tejto nespárenej epitelovej kosti sa na spodnej časti lebky vyvíjajú aj dve párové kosti, nazývané pre-hrot (praevomér); a keďže plaz má otvárač vo forme spárovanej kosti a parasfenoid nebol v tejto oblasti spárovaný s kosťou, za homológa tejto poslednej plazovej kosti je možné považovať cicavčieho otvárača a predozeri sa považujú za otvárače. Predné steny nosnej dutiny sú čeľuste (maxillare), na ktorých sa tvoria škrupiny, rastúce do nosovej dutiny; je to čeľusť (maxilloturbinale).

Predná strana lebky končí vpredu maxilárnymi kosťami (praemaxillare). Táto kosť u antropomorfných opíc a u ľudí rastie veľmi skoro s čeľusťovou kosťou, zatiaľ čo u netopierov sa redukuje na malú kosť alebo vôbec chýba. Maxilárna kosť svojím vertikálnym procesom obmedzuje predný otvor nosa a vstupuje medzi nosnú kosť a čeľusť ďaleko dozadu, niekedy do prednej kosti. Spodné medzimaxilárne kosti sa podieľajú na tvorbe tvrdého patra. Obmedzujú predný a bočný otvor tvrdého podnebia otvorenia podnebia-posovogo (stenson) kanál (pre. Incisiva), spájajúci ústnu dutinu s Jacobsonovým orgánom. Procesy čeľustných kostí ležiacich mimo týchto podnebia-nosových otvorov sa prekrývajú pre každú kosť nezávisle, sú to pravdepodobne homológy párov plazov (praevomérov) a udržiavajú sa v platypuse ako nezávislé kosti. Na maxilárnej kosti sú rezáky.
Čeľuste kosti (maxillare), ako už bolo uvedené, obmedzujú dno nosnej dutiny a vytvárajú tvrdé podnebie horizontálnych procesov. Na vnútornej strane nosovej dutiny k jej bočným stenám, tvoreným čeľusťovými kosťami, rastie spodná opotrebovaná škrupina (maxilloturbinae), položená ako nezávislá kosť.
Pokračovanie tvrdého podnebia sú horizontálne procesy palatínových kostí (palatinum), ktoré obmedzujú vnútorný nosový priechod ako bočné steny. Stoja zvisle a zvisle stojaca pterygoidná kosť (pterygoideum), priliehajúca k ľudskému telu, rastie so sfenoidnou kosťou a tvorí jej pterygoidné procesy (proc. Pterygoidei).
Predná časť lebky prechádza významnými úpravami v súvislosti s prechodom na vodný život u veľrýb, sirén a v súvislosti s vývojom kmeňa slonov.

U veľrýb (obr. 536) sa nosné dierky tlačia ďaleko dozadu do interorbitálnej oblasti a vedú k vertikálnemu kanálu, ktorý otvoria chóry do ústnej dutiny; nosné kosti ležia za nosnými dierkami, spočívajú na predných kostiach a takmer susedia s intertechálnou kosťou; na čelných kostiach ležali aj čeľusťové kosti, siahajúce takmer k parietálnym kostiam, silne posunuté do strán parietálnou kosťou.
Sirény majú rovnaký pohyb zadných nosných dierok, ale nie tak ďaleko; tu sú nosové kosti ešte viac redukované a môžu dokonca chýbať (Obr. 537).

U slonov s nadmerným vývojom kly a silných stoličiek a kvôli prítomnosti kmeňa je normálna poloha kostí silne pozmenená: nosné dierky sa silno tlačia dozadu, preto nosový kanál zaujme takmer kolmú polohu; predné kosti veľmi krátke, maxilárne kosti veľmi veľké a siahajú ďaleko dozadu; čeľusťové kosti posunuté pod základňou lebky; Celá lebka je veľmi masívna s extrémne rozvinutou pneumatickosťou kostí (obr. 538).

Na rozdiel od všetkých ostatných stavovcov sa spodná čeľusť cicavcov na každej strane vyvíja iba z jednej zubnej kosti (dentale); obe polovice alebo zostávajú navzájom spojené pomocou chrupavky, stehu, alebo rastú spolu (monohrypnye, slon, chiropraktici, opice). Každá polovica pozostáva z horizontálneho tela (corpus), na ktorom sú zuby a zo vzostupnej časti s procesmi: predný koronárny proces (proc. Coronoideus), kĺbový proces ležiaci za ním (proc. Articularis) a u dolných cicavcov pomocou uhlového procesu, ktorý sa pohybuje dozadu (proc. angularis); u väčšiny vačnatcov a v jednotlivých hlodavcoch sa tento proces transformuje na dosku siahajúcu smerom dovnútra (obr. 539).

Štruktúra dolnej čeľuste a jej zvláštne uchytenie k lebke nás vedú k otázke pôvodu týchto znakov u cicavcov. Odpoveď na túto otázku poskytuje história individuálneho vývoja a paleontologické údaje. U embryí cicavcov sa kladú štvorcové chrupavky a palatíny. Osifikácia palatínovo-kvadratickej chrupavky sa však vyskytuje mimo križovatky dolnej čeľuste a nevedie k vytvoreniu štvorcovej kosti, ako u plazov, a dá novú zvukovú kosť, incus, zatiaľ čo Meckelova chrupavka, osifikovaná v zadnej časti, dáva neartikulárnu kosť (artikulare), rovnako ako v iných maxilárnych a maxilárnych, a tretia sluchová kosť - malleus. Prvá sluchová kôš, strechy (svorky), zodpovedá stĺpikom obojživelníkov, plazov a vtákov, a ako už vieme, ide o modifikované zavesenie (hyomandibulare), to znamená proximálnu časť hyoidného oblúka. Rovnako ako stĺpec (columella auris), strmeň zakrýva oválne okno zvukového bludiska a je, podobne ako niektoré plazy, prepichnutý dierou, cez ktorú prechádza tvárová tepna (a. Facialis) cez niektoré nižšie cicavce (obr. 540 a 541).

U cicavcov sa teda štvorcová kosť zmenila na nákovu, kĺbovo na malleus; a keďže malá integumentárna kosť, ktorá tvorí stred jej procesu, rastie na malleus, predpokladá sa, že táto integumentárna kosť (goniale) plazov. Epiteliálna uhlová kosť plazov, ležiaca vedľa tejto poslednej kosti, tiež prechádza do ušnej oblasti a stáva sa tympanickou kosťou (tympanikum). Tieto zmeny sú viditeľné na priloženom diagrame (Obr. 541).
Paleontologické údaje potvrdzujú možnosť vyššie uvedených transformácií. Medzi zvieracími nositeľmi zvierat so zubami (Theriodontia) sú také, u ktorých sa zubná kosť stáva obzvlášť veľkým vývinom a dosahuje za spojom dolnej čeľuste štvorcovú kefu. Zároveň šupinatá kosť zostupuje pozdĺž štvorcovej kosti k rovnakému kĺbu. Zubné a šupinaté kosti sa preto čoraz viac zbližujú.
Nakoniec sa u niektorých zubov zvieracích zubov a šupinatých kostí podieľajú na artikulácii spolu s kĺbovými a štvorcovými kosťami.
Čo sa týka sublingválneho aparátu cicavcov, pozostáva z tela, modifikovanej bazálnej časti (kopuly) hyoidného oblúka, predných rohov (cornua anteriora), tvorených spodnou časťou hyoidného oblúka a zadných rohov (cornua posteriora), transformovaných z prvého vetvového oblúka. Predné rohy pozostávajú z niekoľkých osifikácií (až do štyroch); segment, ktorý je obrátený k lebke, často rastie do skalnatej kosti, ktorá sa tvorí styloidný proces (proc. styloideus).
Druhý a tretí vetvový oblúk spôsobil vznik chrupavky štítnej žľazy (chrupavka tyreoidea.)
Opasky končatín. Ramenný opasok cicavcov sa vyznačuje predovšetkým znížením kostí kostí (procoracoideum, epicoracoideum, coracoideum), tendenciou vymiznúť kľúčnej kosti (clavicula) a dobre rozvinutým hrebeňom lopatky (spina scapulae), ktorý pokračuje v akromionovom procese (acromion).
Avšak, jeden priechod ramenný opasok má veľkú podobnosť s plazmi. Lopatka je opatrená akromionárnym výstupkom, na ktorom leží kľúčnica v prednej časti, a na lopatke sú pripevnené široké a krátke kosti havranov (coracoideum), siahajúce až k hrudnej kosti. Pred nimi sú široké prokokaidy (procoracoideum) vo forme nezávislých kostí. Klavikuly (clavicula) sa mediálne spájajú s epistémom (episternum) (Obr. 542). U ostatných cicavcov sa kukidy kladú vo forme dvoch osifikácií, strácajú kontakt so sternom a priľnú k lopatke, čím sa vytvorí kokosový proces (proc. Coracoideus); procoracoids úplne zmiznú (Obr. 543), v niektorých dravých jedincoch a kôstkách zmiznú v spojitosti s adaptáciou na rýchly beh a u veľrybotvarých a sirén v súvislosti s transformáciou predných končatín na plutvy.


Panvový pás cicavcov má na úchytke typické usporiadanie prvkov; jeho vlastnosťou je, po prvé, prítomnosť ďalšej kosti v acetabula (acetabulare), po druhé, fúzia všetkých týchto kostí na každej strane do jednej bezmennej kosti (innominatum) a fúzia medzi ochlpeniami na ochlpenie a sedacie kosti, takže uzavretá všetky strany otvoru medzi týmito kosťami (pre. obturatum) (Obr. 544). V monotrómy a vačnatých tkaninách sa na ochlpenie ochlpenia (marsupiale) pripája špeciálna kosť.
Kostra končatiny v pôvodných formách cicavcov je to kvôli ich prispôsobeniu rýchlym pohybom - beh pomocou štyroch končatín, ktoré sa nachádzajú pod telom, takže ten nevisí medzi končatinami, ako väčšina plazov, ale spolieha sa priamo na ne. Schopnosť bežať je spôsobená odlišnou polohou končatín u cicavcov. Vo svojich končatinách sú orientované tak, aby lakťové kĺby predných končatín smerovali dozadu a kolenné kĺby zadných končatín smerovali dopredu. Pri pohybe sú predné končatiny vyhodené dopredu a pritiahnutím tela priliehajú k telu lakťové kĺby; zadné končatiny tlačia trup dopredu narovnaním nôh pri kolennom kĺbe.

Podľa počtu končatín, ktoré tvoria kosti a ich umiestnenie, má kostra končatín prvotných foriem cicavcov veľmi typickú štruktúru primitívnej päť končatiny a v rámci triedy prešla veľkými zmenami iba v procese adaptácie na rôzne druhy pohybov v rôznych podmienkach prostredia.
V prednej končatine sa vyznačuje prítomnosťou v distálnej časti ramennej otvory (pre. entepicondylare), prítomnosť proximálneho výčnelku ulna kosť (olecranon), prítomnosť hrachu v tvare hrachu (pisiforme) a fúzia štvrtého a piateho distálneho semena karpálneho semena (Carpalia dis talia 4 + 6 = hamatum alebo uncinatum) a centrálnej proximálnej k intermediálnej (centrálnej dist. + intermedium = lunatum alebo semilunare). B vzhľadom na to, že sa často používa nomenklatúra osciftu zápästia cicavcov, prijatého v ľudskej anatómii, uvádzame tabuľku týchto mien v diagrame označujúcom umiestnenie kostí.
Zadnú končatinu cicavcov charakterizuje predovšetkým skutočnosť, že kolenný kĺb pred stehnom má dobre vyvinuté procesy na proximálnej časti - trochanter 2-3, trochanter major, minor a terius; v holennej kosti je holenná kosť rozvinutejšia, je tu kosť jabĺčko (patella), chráni kolenný kĺb. V dechte sú iba dva proximálne kosti, dobre vyvinuté: vnútorný - talu (talu = astragalus), vytvorený zo sútoku stredného a stredného proximálu (intermedium + centrale proximale) a vonkajšej päty (calcaneus), ktorá zodpovedá vláknitej kosti preplute (fibulare). Zo semien v strednej rade sa stredná distálna (centrálna distálna) zmenila na loďovitý tvar (naviculare = scaphoideum). Tibiálna kosť bola buď zredukovaná alebo k nej pripojená.
Usporiadanie semien tarsu a tarzu a synonymá mien sú takéto:
V súvislosti s prispôsobením sa pohybu v rôznych prostrediach končatiny cicavcov prešli veľmi významnými zmenami a tieto zmeny je možné obzvlášť dobre sledovať na kostre. Bohužiaľ nemáme žiadne paleontologické údaje o štruktúre končatín predkov cicavcov. Nie je pochýb o tom, že prvé cicavce mali malé špecializované končatiny vyššie uvedeného typu. Takéto končatiny moderných cicavcov majú najprimitívnejšie vačnatce, napríklad vačice - vačnaté potkany (Didelphys) a hmyzožravé, čo vedie k drevnatému spôsobu života. Z toho by však bolo nesprávne vyvodiť, že predkovia cicavcov boli v každom prípade lesnými obyvateľmi a okrem horolezeckých foriem. Okrem toho končatina vačice, hoci slúži na pohyb po uzloch stromov, nie je v žiadnom prípade typickou špecializovanou končatinou, ale zachováva si svoj univerzálny päťprstý typ. Takéto končatiny by sa dali najlepšie uchovať na zvieratách, nielen na zemi, ale aj na lezenie na stromoch, ako aj na zvieratách, ktoré lezú na hory. Špecializácia končatín cicavcov prebiehala súčasne s diferenciáciou rádov a ďalších menších systematických jednotiek, pričom došlo k zbližovaniu.
Končatiny sa podrobujú obzvlášť hlbokým úpravám pri prispôsobovaní sa letu, plávaniu vo vode, pohybu pod zemou; pomalé pohyby po zemi a život stromu za to nestoja kvôli hlbokej špecializácii.
U netopierov (Chiroptera) sa predná končatina zmení na krídlo, pričom letiaca membrána sa tiahne medzi štyrmi (2-b) vysoko pretiahnutými prstami, bez chĺpkov; prvý prst je krátky a na konci má silne vyvinutú pazúr; tento prst nehrá pri lietaní žiadnu úlohu, slúži na lezenie; v zápästí spolu vyrastie semifinále lunárne a stredná kosť v tvare lode; predlaktie je pretiahnuté a pozostáva z dobre vyvinutého polomerzatiaľ čo ulna je znížená a rastie na polomer. Lietajúca membrána zachytáva a zadné nohy. Medzi nimi je vyvíjaný pre netopiere inak (Obr. 545).
U cicavcov plávajúcich vo vode sa končatiny menia na plutvy rovnakým spôsobom, ako sme už videli u plazov. Obzvlášť sa zmenili končatiny veľrýb a sirén. Na rozdiel od krídla je predná končatina v jej proximálnej časti výrazne skrátená. Vo veľrybách sa kosti ramien a predlaktia zmenili na krátke a široké platne, čo dáva priestor na pripevnenie silných svalov. Všetky časti končatín sú navzájom pevne spojené a tvoria silné lopatky. Zároveň sa môže výrazne zvýšiť počet falang. Rovnaké zmeny sme videli aj u plazov. Nárast počtu falang závisí od faktu, že diafýza falangy nerastie spolu s epifýzami a osifikuje z jednotlivých centier. Takáto samostatná osifikácia je u týchto zvierat spojená s pomalým procesom osifikácie. Spomalenie procesu osifikácie vedie k tomu, že väčšina kostí v kĺboch ich zvyškov chrupavky.
Plutvy Siren (Sirenia) sú podobné ako u veľrýb veľrýb: podobajú sa navzájom: rameno a predlaktie sú kĺbovo spojené, aby sa mohli ohýbať; sú takmer všetky kosti zápästia a všetkých päť prstov s mierne zmeneným počtom falang (obr. 546).

Zadné končatiny moderných veľrýb a sirén sú zmenšené do tej miery, že z nich zostávajú iba zvyšky panvy (Obr. 547 a 548).
Ďalší typ adaptácie končatín na pohyb v krbu je vidieť v Pinnipedia. Chvost takmer zmizol a zadné nohy sa pohybujú dozadu a hrajú úlohu motorického orgánu. Tri rodiny tohto rádu sú zastúpené tromi stupňami (nie geneticky) podľa stupňa rozvoja adaptácie na pohyb vo vode. V mrožiach a ušiach ušatých (Otariidae) sa zadné končatiny môžu stále ohýbať vpred pod telom, ale v mrožiach sú v limbickom zariadení primitívnejšie zariadenia, sú kratšie a pohyblivejšie. V tuleňoch (Phocidae) sa zadné končatiny nemôžu ďalej ohýbať pod telom dopredu, ale pazúry stále zostávajú na prstoch (plutvy).
U drtiacich zvierat sa končatiny tiež veľmi silne modifikujú: končatina molice vačnatej sa zmenila najmä (Typory hlodavcov Notoryctes). Má dva hrebene na pleci, podobne ako mnoho nórskych zvierat, čo je spôsobené silným vývojom tricepsového svalu; existuje kľúčná kosť, rovnako ako všetky kopajúce cicavce. Na humeru je veľmi silný hrebeň, vidlička na vidlice a slúži na pripevnenie silného (potrebné pri vykopávaní) deltoidu a prsný sval; vnútorný kondyl ramena je silne vyvinutý vďaka silnému rozvoju flexorových svalov ulnárnej kosti (proc. olecranon), ktorá slúži ako miesto pripojenia tricepsov; zápästie sa špeciálne mení, stáva sa veľmi odolným; 1. a 2. prsty sú zasunuté na strane dlane a iba 3. a 4. prsty sa používajú na kopanie, vybavené obrovskými pazúrmi, najmä tretím; počet falang v prstoch je veľmi malý: 2. a 1. 5. metakarpale so širokou plochou pazúrkou, ktorá zväčšuje kopací povrch; v rovine dlane je sesamoidná kosť, ku ktorej je sval pripojený, ohýbajúc prsty, ktoré hrajú veľkú úlohu pri kopaní; To zvyšuje pevnosť zápästia; podobne ako iné formy kopania, je symfýza ohniskovej kosti veľmi krátka; ischiálna kosť rastie spolu s priečnymi procesmi dvoch posledných sakrálnych stavcov; na stehne, kolene a holennej kosti sú hrebene a výrastky - miesta pripojenia svalov - dobre vyvinuté; kosti holene a fibuly (holene a fibuly) pevne spojené, rovnako ako kosti zlatých rýb (Chrysochloris); podrážka smerom von, napríklad Chrysochloris; Piaty prst je ohnutý na plantárnom povrchu, piaty klinec na chodidle je široký a špachtľový, štvrtý je dlhý a kosákovitý, tretí a druhý sa zmenšujú a rozširujú; na hrudnej kosti silne vyvinutý hrebeň, podobne ako krtko zlatých listov, krtek krtko (Spalax) a krtek (Talpa) (obr. 549).
Silný vývoj hrebeňov na kostiach na prichytenie svalov, skrátenie karpusu, pevné spojenie karpálie, zníženie počtu falang a posilnenie iných, ktoré zohrávajú úlohu pri kopaní - všetky tieto znaky sú všeobecne charakteristické pre kopanie cicavcov. Ale v závislosti od pôdy, v ktorej zviera kope, sa pozorujú rozdiely. Pri kopaní do mäkkej zeme (napríklad Talpa v móle) (obr. 550) sa kefa veľmi rozširuje, pôsobí ako lopata a má tendenciu vyvíjať šiesty prst. Predná časť krtka je porovnateľná s lopatou používanou na kopanie v mäkkej pôde. Naopak, pre tie formy, ktoré kopajú v tvrdom teréne, sa kefa stáva užšou, iba pár prstov sa zúčastňuje kopania, z väčšej časti 3. a 4., ktoré sú silnejšie ako iné, napríklad v Notoryctes a Chrysochloris. Táto kefa je porovnateľná s rýľom na kopanie v tvrdom teréne.
![]()
To isté vidíme u cicavcov, ktorí trhajú solídne konštrukcie termitov, napríklad v mravcovi (Myrmecophagus), v tamandue. Tieto zvieratá chodia, opierajúc sa o vonkajší okraj chodidiel a ohýbajú svoje pazúry.
Adaptácie cicavcov na rýchly pohyb po zemi spôsobili veľmi významné zmeny končatín. Pohyb na zemi, ktorý bol charakteristický pre primitívne cicavce, sa uskutočnil spoliehaním sa na celú ruku a nohu; Počiatočné formy cicavcov boli predovšetkým zastavujúce alebo planételové, to znamená, že pri chôdzi sa spoliehali na celú nohu, napríklad na medvede. Takáto štruktúra však umožňuje len relatívne pomalú chôdzu v porovnaní s antilopami alebo koňmi s ušami, alebo skákajúcimi zajacami, klokanmi, jerboami. Počas fylogenetického vývoja vidíme postupné zdvíhanie zápästia a trupu (carpus a tarsus) nad zemou, po ktorom nasleduje metakarpus a tarzus, ale falangy sa stále úplne dotýkajú zeme. Nakoniec sa falangy narovnali a predné a zadné končatiny začali spočívať na koncoch prstov. Štádiá tohto vývoja môžeme teraz krok za krokom sledovať nielen u kopytníkov, ale aj u mäsožravých cicavcov, ktorí získali schopnosť rýchlo bežať. Keď sa končatiny narovnávajú, extrémne prsty sa už nedotýkajú zeme, prestávajú fungovať a podliehajú atrofii. Ukázalo sa, že povrchový kontakt je menší. Zmeny v distálnej časti končatín znamenali zmeny v proximálnej časti: karpália a tarzália sú umiestnené bližšie vedľa seba, ulna a fibula sú znížené, končatiny začínajú vykonávať pohyby iba v sagitálnej rovine, stredná metapodia sa spája do jednej kosti.
Dve cesty vývoja vedú k formovaniu končatín moderných kopytníkov. V jednom riadku prechádza os končatiny uprostred tretieho prsta a posledný sa vyvinie najsilnejšie, zvyšok sa stáva základným. Sú to tuhé látky (Perissodactyla), mezaxiálne (Mesaxonia). U ostatných os prechádza medzi tretím a štvrtým prstom, ktoré sa vyvíjajú rovnomerne: parciálne (Artiodactyla), paraxiálne (Paraxonia). Bočné prsty sú zmenšené. Vývoj pevných látok je najzreteľnejšie viditeľný vo fylogenetickom vývoji končatín (Obr. 551). Vývoj kopytnatého štátu bol okrem iného spôsobený tým, že mu predchádzalo zníženie prvého prsta, takže pri prechode zo zastavenia sa takmer nezúčastnil, zatiaľ čo u predkov monokrytov bol vývoj piateho prsta pôvodne normálny,
Je zaujímavé, že v evolučnom procese sa objavili parochidry aj objavenie opakovane a v niektorých prípadoch viedli k veľmi špecializovanému typu, napríklad v Južnej Amerike (zaniknutý).
Ostatné prsty sú pokryté kopytami, ktoré sa môžu ďalej odlíšiť v závislosti od životného štýlu a lokality. Kozy, ktoré držia skaly, kopyta, sú teda strmé a úzke a vybavené ostrou hranou; v soboch sú veľmi dlhé, široko rozdvojené a slúžia ako druh lyží, ktoré zabraňujú tomu, aby zviera padalo na sneh a močiare.
Súčasne s prstami počas vývoja chôdze po prstoch dochádza k zmene zápästia (karpusu) a tarzu (tarzu). Pre artiodaktyly a tuhé látky to platí inak v dôsledku rozdielneho rozdelenia gravitácie. Nakoniec, v súvislosti s prechodom na nalcezhhozhdeniyu postupne znižuje hodnotu ulna (kosti) ako podporu pre rameno: znižuje alebo sa spája s polomerom (polomer). Rovnakým spôsobom sa zníži fibula a nakoniec zmizne v zadných končatinách.
Význam a povaha opísaných zmien sú jasne viditeľné z porovnania končatín ťava a slona (obr. 552). U slona zostáva ruka (noha) krátka, zatiaľ čo rameno a predlaktie (stehno a holenná časť) sú predĺžené. Naopak, ťava predlžuje rastúce metapódie; horná časť paže a predlaktie (rovnako ako stehno a dolná časť nohy) zostávajú krátke. V dôsledku straty množstva kostí dochádza k zjednodušeniu kostry končatiny, ktorá sa stáva ľahšou. Redukcia prstov vedie k menšiemu treniu, k zníženiu rizika poškodenia, prasknutiu väzov. Akreácia metapódií vytvára väčšiu silu. Končatina slona je oveľa ťažšia. Hojnosť kostí v ruke vyžaduje dosiahnutie stability spojenia všetkých prstov s bežnou pokožkou. Končatina slona je stĺpec veľkej sily, ale chýba mu ľahkosť končatiny ťavej končatiny.
Ďalším spôsobom, ako sa rýchlo pohybovať po zemi, je skákanie. Táto adaptácia sa objavila opakovane v histórii cicavcov a naviac v širokej palete fylogenetických kmeňov: v sérii vačnatcov na klokanoch (Macropus), medzi hmyzožravcami v mikinách (Macroscelidae) a nakoniec v rôznych radoch vývoja hlodavcov: sem. zajace (Leporidae) z Duplicidentata; v rade ošípaných (Hystricomorplia) sa vyvinula skupina dlho korene (Pedetinae); v rade miciel (Myomorpha) máme austrálsky jerboa (Conilurus), jerbay starého sveta (Jaculus, Alactaga), americké jumpery (Zapus) a vreckové jumpery (Dipodomys).
Rozvoj schopnosti skákať sa spája s predlžovaním zadnej končatiny, konkrétne kostí metatarzu (metatarzu). Okrem toho, keď k tomu dôjde, strata nepotrebných extrémnych prstov a kosti tarzu sú navzájom pevne spojené. Príkladom by mohli byť nožičky zajaca zeme (Alactaga) a skutočné jerboi (Dipus).
Predné nohy Alactaga sú malé a krátke. Zajac ich používa pri kopaní, ale pri skoku sa nedotýka zeme. Skákanie sa vykonáva iba pomocou zadných nôh a dokonca aj bez chvosta. Ten sa dotýka zeme iba v pokojnej polohe, keď zviera spočíva na chvoste. Kostra chodidla má zvláštnosť, že metatarzálne kosti 2., 3. a 4. prsta sa zlúčia do jednej kosti. Konce metatarzálnych kostí dole nie sú spojené a stredný (tretí prst) vyčnieva ďalej vpred. Z falošných spojov, ktoré sú s nimi spojené, sú opäť tie najdlhšie. 1. a 5. prst sú základné a nedotýkajú sa zeme. Obidva tieto prsty majú rovnakú dĺžku v dôsledku skutočnosti, že metatarzálna kosť palec dlhšie ako 5. Prvý má 2, druhý - 3 falangy.
Špecializácia Dipus išla ešte ďalej: 1. prst zmizol bez stopy, od piateho sa zachoval iba základ. Prostredný prst dvakrát slabší ako dva extrakty a iba o niečo dlhšie ako dva. Prsty sú paralelné. Prostredný prst je zmenšený, pretože 2. a 4. preberajú svoju funkciu, ktorej konce kladú dôraz na sedenie a skákanie. V ďalšom vývoji by mal tretí prst zmiznúť: v Dipuse máme neúplný typ adaptácie.
Vonkajšia štruktúra psa
Pes bol prvým miláčikom. Muž to skrotil v staroveku. Pes počas lovu pomáhal primitívnemu človeku, strážil jeho domov. Teraz známe služobné psy, poľovnícke a okrasné, ako aj mutty (čistokrvné). Psi pomáhajú vykonávať službu stráženia hraníc našej krajiny a počas Veľkej vlasteneckej vojny našli zranených, pomohli v spravodajských službách a komunikáciách. Vedci uskutočňujú na psoch štúdie tela. Pes býval vo vesmíre.
Telo psa, ako je pastiersky pes alebo husky, je štíhle a svalnaté. Končatiny sa nenachádzajú na bokoch tela, ako u plazov, ale pod telom, takže telo zvieraťa sa nedotýka zeme. Pes kráča a opiera sa o prsty so silnými pazúrmi. Flexibilný krk prispieva k väčšej pohyblivosti hlavy. Ústa zvieraťa sú obmedzené pohyblivými perami - hornými a dolnými. Nad hornou perou je nos s párom vonkajších nosných otvorov - nosných dierok. Oči majú dobre vyvinuté očné viečka. Blikajúca membrána (tretie očné viečko) u psa, rovnako ako u všetkých cicavcov, je nedostatočne rozvinutá. Zo všetkých zvierat majú iba cicavce vonkajšie ucho - ušnica. Vonkajšie ucho psa je veľké a pohyblivé.
Cicavčie čísla
Koža cicavcov je silná a elastická: u väčšiny zvierat sú do nej umiestnené základne vlasov, ktoré tvoria charakteristiku tejto triedy stavovcov. vlasový kabát, Sú silné a dlhé vlasy - osina a kratšie, mäkšie - pod tlakomalebo základný náter, Drsná a odolná markíza chráni spodnú vrstvu a pokožku pred poškodením. Podsada, ktorá zadržuje veľa vzduchu, dobre udržuje telesné teplo. Okrem markízy a spodku sa u zvierat vyvinú veľké vlasy - orgány dotyku.
Cicavce z cicavcov, podobne ako perie vtákov a šupiny plazov, pozostávajú z rohovky. Dvakrát do roka pes, rovnako ako mnoho iných cicavcov, prístrešok - časť jeho vlasov vypadne a je nahradená novými. Dno vlasov sa nachádza vo vnútri špeciálneho vlasového vrecka, kde sa otvárajú kanály priľahlých mazových žliaz. Ich sekréty mažú pokožku a vlasy, vďaka čomu sú elastické a nie sú navlhčené vodou.
Obrázok: Štruktúra kože a vlasov cicavca
V koži sa väčšina zvierat nachádza a potné žľazy, Pot, ktorý sa vyparuje z povrchu tela, ho ochladzuje. Spolu s potom sa z tela odstráni aj nadbytočná soľ a močovina; potné žľazy teda zohrávajú úlohu ďalších vylučovacích orgánov.
Pes má málo potných žliaz a ochladzovanie tela sa dosahuje zvýšeným dýchaním.
Na koncoch cicavčích prstov sú rohové pazúry, klince alebo kopytá. Formácie rohov sa niekedy vyvíjajú aj na hlave (rohy nosorožcov, antilopy, hovädzí dobytok atď.) Alebo na chvoste (napríklad rohovité šupiny u potkanov).
Cicavčia kostra
Kostra cicavcov pozostáva z rovnakých častí ako u ostatných stavovcov. Lebka zvierat sa líši vo väčšej lebke, ktorá je spojená s väčšou veľkosťou mozgu. Pre cicavce je veľmi charakteristická prítomnosť 7 krčných stavcov. Žirafa s dlhým hrdlom a veľryby majú rovnaký počet krčných stavcov. Hrudné stavce (zvyčajne 12 - 15) spolu s rebrami a hrudnou kosťou tvoria pevnú hruď. Masívne stavce bedrový pohyblivo kĺbovo medzi sebou spojené. V tejto časti sa trup môže ohýbať a uvoľňovať. Počet bedrových stavcov sa u rôznych druhov líši (2-9), u psov je ich 6. Sakrálna chrbtica (3-4 stavce) rastie spolu s kosťami panvy. Počet stavcov kaudálnej časti (od troch do niekoľkých desiatok) závisí od dĺžky chvosta.
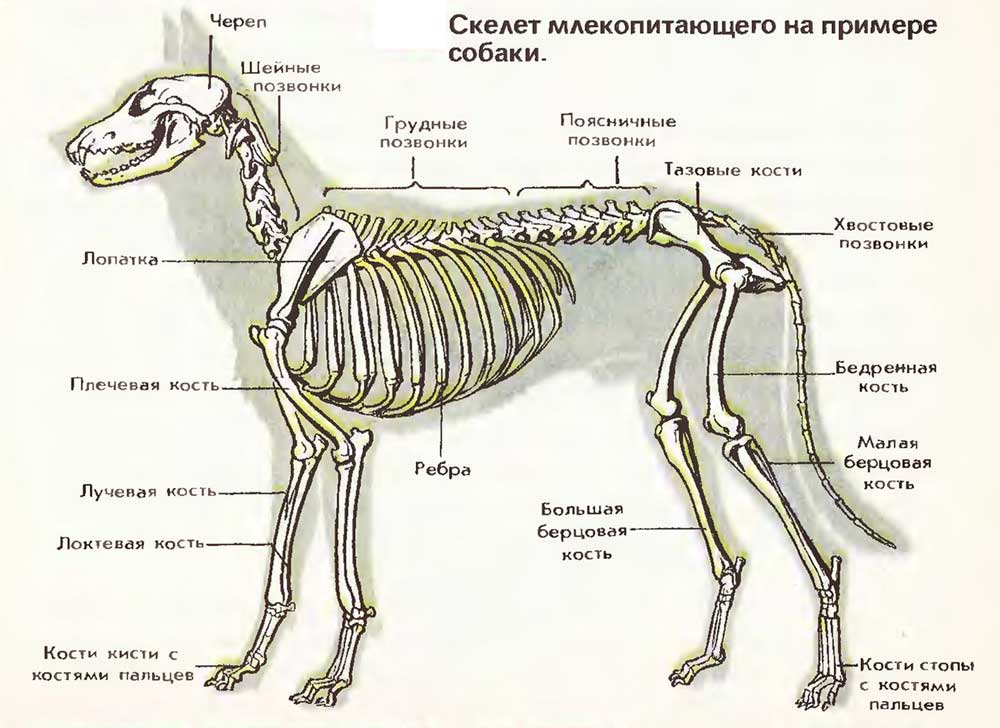
Obrázok: Kostra cicavcov na príklade psa

Obrázok: Pes Lebka
Predný ramenný pás Cicavce sa skladajú z dvoch čepelí, ktoré sú k nim pripevnené vranami, a dvoch klinčekov. Pes nemá kosť. Zadný pás - panva - tvorené tromi pármi panvové kosti, Kostry končatín u rôznych druhov cicavcov a plazov sú podobné, ale podrobnosti o ich štruktúre sa u rôznych druhov líšia a závisia od biotopu zvieraťa.
Svalové cicavce
U väčšiny cicavcov vrátane psa sú zvlášť vyvinuté svaly chrbta, končatiny a ich pásy. Pes môže bežať vo veľkých skokoch, ohýbať a narovnávať trup, striedavo tlačiť zo zeme prednými alebo zadnými nohami. Silné svaly sú v pohybe. dolná čeľusť, Zaťaté čeľuste, vyzbrojené zubami, pes pevne drží korisť.

Obrázok: Svaly psov pohybujúce sa spodnou čeľusťou