§ 9. Череп
Филогенетические преобразования . Филогенез черепа может быть прослежен при изучении сравнительно-анатомических и палеозоологических данных.
Уже у примитивных хрящевых рыб (акул), как и у всех позвоночных, череп резко разделен на два отдела - мозговой и висцеральный (рис. 40, А). Мозговой череп вмещает головной мозг, парные органы слуха и обоняния и образует боковые выемки, в которых помещаются глазные яблоки. К мозговому черепу подвижно прикрепляется висцеральный, или внутренностный (viscera - внутренности), череп. Это прообраз лицевого черепа. Он охватывает ротовую полость и глотку с жабрами. Висцеральный череп состоит из ряда парных - висцеральных - дуг. Впереди лежит челюстная, за ней подъязычная, затем несколько жаберных дуг. Челюстная дуга на каждой стороне образована двумя сочленяющимися друг с другом хрящами: нёбно-квадратным (сверху) и нижнечелюстным. Подъязычная дуга тоже состоит из двух члеников: подъязычно-челюстного и подъязычного хрящей. Первый из них верхним концом сочленяется с черепной коробкой и дает опору челюстной дуге, а нижним - с подъязычным хрящом. Жаберные дуги состоят из нескольких хрящей каждая.
Новорожденные щенки плаценты находятся на гораздо более высоком уровне развития, в отличие от сумчатых, и многие из них способны к активной жизни сразу после рождения. Важные особенности плаценты включают увеличенный череп, хорошо развитый костный пластырь, точное количество позвонков и расщепление ребер. Основными формулами плацентарных протезов являются 3 резца, 1 собака, 4 премоляра и 3 моляра. Очевидно, что некоторые плаценты были результатом специализации определенных отклонений от этого основного деления.
Для большинства животных резцы предназначены для отрезания пищи. Аналогично, функция клыков заключается в том, чтобы кормить пищу. Премоллары обычно имеют сложную конструкцию короны. Молекулы служат отношениям познания. Это зубы с короной, которая имеет сложную структуру.
Череп млекопитающих и человека сплошь костный, остатки хряща у взрослых сохраняются лишь в носовой перегородке.
Число костей значительно сократилось за счет срастания их друг с другом (рис. 41). Верхние челюсти образованы только покровными костями, возникающими уже у костных рыб. Межчелюстные (резцовые) кости закладываются самостоятельно, но вскоре сливаются с верхнечелюстными костями. Нижняя челюсть непосредственно (а не через квадратную кость) сочленяется с височной костью, что составляет специфическую особенность млекопитающих. Квадратная кость превращается в слуховую косточку - наковальню, а остаток нижнечелюстного хряща (сочленовная кость рыб) - в молоточек. Верхняя часть подъязычной дуги уже у амфибий образовала стремя. Таким образом, все три слуховые косточки - стремя, наковальня и молоточек - происходят из остатков висцеральных (челюстной и подъязычной) дуг. Средняя часть подъязычной дуги превращается в шиловидный отросток височной кости, соединенный связкой с нижней частью этой дуги, которая вместе с остатком первой жаберной дуги преобразуется в подъязычную кость. Вторая и третья жаберные дуги образуют щитовидный хрящ гортани; четвертая дуга - дужку перстневидного хряща гортани; пятая дуга полностью утрачивается.
В течение третичного и четвертичного периодов, более 65 миллионов лет, появилось больше плацентарных, некоторые из которых вымерли. На сегодняшний день сохранилось только 16 заказов. В таблице в приложении дается обзор всех известных плацентарных заказов.
Эти млекопитающие, вероятно, являются предками всех других плаценталей. Недавние исследования показывают, что можно было бы разделить все насекомоядные заказы на четыре большие группы. Летучие мыши, жуки и приматы также принадлежат к насекомоядным. Последняя группа Ферунгулата включает зверей и всех копытных, как они вымерли. Процесс плацентарной дивергенции начался во время мела. Появление современных насекомоядных водорослей похоже на появление насекомоядных в мезозое.
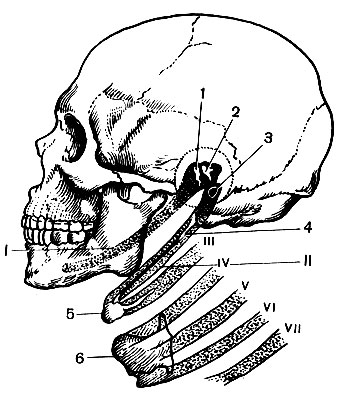
У млекопитающих мозговой отдел черепа обычно значительно меньше лицевого. У человека мозговой отдел более развит и резко преобладает над лицевым (рис. 42).
Мозговой отдел образован непарными костями (затылочной, клиновидной, лобной, решетчатой) и парными (теменными и височными) (Атл., 14, 15, 28). Некоторые кости (клиновидная, решетчатая и др.), расположенные на границе мозгового и лицевого отделов, функционально участвуют и в формировании лицевого.
Они всегда были маленькими животными, и, вероятно, их размер и образ жизни позволили им выжить. У убийц насекомых есть небольшой и относительно простой мозг, изначально состоящий из пяти лимбитов и простой зубной ряд. Наиболее распространенными видами насекомоядных были Дельтатериум, небольшое животное с черепом длиной около 5 см и очень примитивная структура стула.
Построение их тел подобно насекомоподобным. Полет развился уже в эоцене. В отличие от птиц, они разработали совершенно другой способ полета, расширив передние конечности между пальцами мембраны. Сегодня нет никаких сомнений в том, что предки самолета были плоскими. Но мы не знаем какой-либо переходной формы между их предками и обычно разработанными самолетами. На палеоцене нет ископаемых находок, когда эти переходные формы должны существовать.
Теменные кости (ossa parietalia) почти четырехугольны, замыкают череп сверху и с боков. Выпуклые части их называют теменными буграми .
Лобная кость (os frontale) примыкает к переднему краю теменных костей. Она состоит из чешуи, глазничных и носовой частей (Атл., 14, 15). На ее выпуклой чешуе спереди выступают два лобных бугра ; ниже их лежат надбровные дуги , латерально оканчивающиеся скуловыми отростками , а еще ниже находятся две надглазничные вырезки , реже отверстия . На нижней вогнутой поверхности глазничной части , у скулового отростка, расположена ямка слезной железы , а медиально заметна блоковая ямка , иногда шип - это место прикрепления хрящевого блока, через который перекидывается одна из глазных мышц. Между глазничными частями располагается носовая часть, охватывающая решетчатую вырезку . В толще лобной кости находится лобная пазуха , сообщающаяся с носовой полостью.
В настоящее время самолеты содержат 17 семей, 178 родов, а число видов составляет около тысячи. Самолеты обладают исключительной способностью перемещаться в пространстве, передавая и получая ультразвук, позволяя им избегать препятствий и охотиться на насекомых. Глазные органы хорошо развиты.
Единственными продовольственными конкурентами самолетов являются ночные птицы. Поэтому многие виды летучих мышей не только питаются летающими насекомыми, но также нуждаются в разных видах пищи. Некоторые питаются кровью животных, другими растительными продуктами, такими как фрукты, нектар, пыльца.
Затылочная кость (os occipitale) участвует в образовании основания и свода мозгового черепа, который она замыкает сзади и снизу (рис. 43). Кость состоит из вогнутой чешуи , парных боковых частей с яремными отростками и мыщелками (сочленяются с атлантом) и основной части . Эти четыре части ограничивают большое затылочное отверстие . Основание каждого мыщелка пронизано коротким каналом подъязычного нерва . Латерально от мыщелков выдаются яремные отростки . Поперек наружной поверхности чешуи тянутся шероховатые верхняя и нижняя выйные линии и выступает наружный затылочный выступ . На мозговой поверхности чешуи возвышается внутренний затылочный выступ , от которого расходится крестообразное возвышение с широкими бороздами от венозных пазух.
Самые старые определенные останки самолета известны из среднего эоцена. Подчиненные калоны, в отличие от летучих мышей, больше животных, которые питаются исключительно фруктами и живут в субтропиках Старого Света. Подводные летучие мыши распространены по всему миру.
При строительстве зубов и конечностей они остаются очень примитивными плацентами, непосредственно связанными с насекомыми-вредителями. Первые известные приматы вряд ли узнаваемы от насекомоядных. Среди плаценталей мезозоя, вероятно, были и арборикозные формы. Приматы являются одной из групп, которые сохранили этот образ жизни в своем дальнейшем развитии. Расщепление приматов от насекомоядных, вероятно, происходило в нижнем палеоцене или уже в верхнем меле около 80 миллионов лет назад.
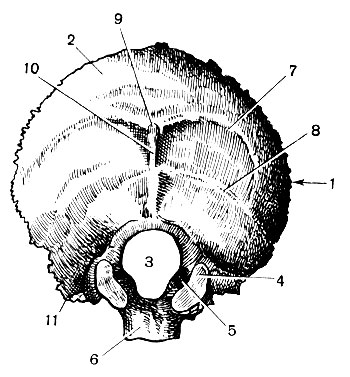
Височные кости (ossa temporalia) примыкают к затылочной. Они участвуют в образовании боковой стенки и основания мозгового черепа, служат вместилищем органов слуха и равновесия, местом прикрепления жевательных мышц и мышц шеи, сочленяются с нижней челюстью. В связи с многообразием функций височная кость имеет сложное строение. На ее латеральной поверхности находится наружное слуховое отверстие , вокруг которого располагаются: сверху чешуя, сзади - сосцевидная часть, спереди и снизу - барабанная часть и медиально - пирамида, или каменистая часть.
Это небольшая плацента с относительно длинным носом и длинным хвостом, который питает насекомых. Его позиция не однозначна по своему характеру. Некоторые эксперты оценивают его на стороне насекомоядных видов, другие - среди примитивных приматов. Некоторые персонажи соответствуют насекомым, другие - приматам и поэтому составляют переходную связь между двумя порядками. Исследования развития приматов сталкиваются с рядом трудностей. Самые старые известные полуопические находки относятся к среднему палеоцене в Северной Америке.
Они показывают ряд специализаций, которые ясно показывают свое раннее происхождение. От этих предков, таких как кожевники, адаптивное излучение начало развивать приматов в различные линии. Сегодня мы признаем три основные группы. Из палеоцена и эоцена мы знаем относительно много семей приматов. Однако обе группы исчезают в конце эоцена, и из олигоцена известно лишь несколько остатков. Только в миоцене происходит новое развитие приматов, в которых доминирует антропоидная группа. В эту группу входят современные лемуры.
Чешуя - слабовогнутая пластинка, замыкающая мозговой череп сбоку. На ней выдается обращенный вперед скуловой отросток , соединяющийся со скуловой костью. Под его основанием находятся сочленовные ямка и бугорок. Здесь происходит сочленение с головкой нижней челюсти.
Сосцевидную часть образует сосцевидный отросток (место прикрепления мышц), легко прощупываемый через кожу за ушной раковиной. Внутри отросток состоит из небольших воздухоносных полостей - ячеек. В отличие от других пневматизированных костей они сообщаются с полостью среднего уха.
Однако нельзя отмечать обезьян палеоцена и эоцена как лемурические, если у нас нет более глубоких знаний. Не исключено, что некоторые семьи получают своих персонажей до конца палеогена. Первые полуполы появляются на Среднем палеоцене Северной Америки. Однако филогенетически эти формы заканчиваются без потомства. После вышеупомянутого первого излучения приматов сле - дует другое излучение в эоцене. Во время этого относительно длительного излучения сформировалась группа приматов антропоида, которая включает ели, макаки, гиббоны, орангутаны, шимпанзе и человека.
Барабанная часть меньше других частей, она ограничивает наружный слуховой проход.
Пирамида , или каменистая часть , заключает в себе барабанную полость и полость внутреннего уха. На ее задней поверхности расположено внутреннее слуховое отверстие , а латеральнее его - щелевидное отверстие водопровода преддверия . На передней поверхности заметна плоская крыша барабанной полости и медиальнее ее - дугообразное возвышение . На вершине пирамиды находится небольшая ямка узла тройничного нерва. На нижней поверхности выступает шиловидный отросток и находится наружное отверстие канала сонной артерии . Этот канал проходит внутри пирамиды и открывается затем на ее вершине одноименным отверстием. Между шиловидным и сосцевидным отростками расположено шило-сосцевидное отверстие канала лицевого нерва. В углу между чешуей и пирамидой открывается мышечно-трубный канал , заключающий в себе слуховую трубу, ведущую в полость среднего уха.
Ближайшим живым родственником человека является шимпанзе. Относительность подтверждается генетическими исследованиями, а степень генетического разнообразия аналогична степени близких видов. Систематически это семейство оценивается по-разному и доминирует над двумя взглядами. В систематической классификации высших обезьян нет единообразия.
Начало развития зубного протеза и вертикальная ходьба должны быть приведены в связь с переходом от леса к саванне. Новая среда, вероятно, способствовала следующим анатомическим изменениям. Именование персонажа, который может быть показан на костях конечностей и изменения в области таза и позвоночника. В то же время существует другая разработка передних конечностей, выпущенных для выполнения других задач. Новые экологические стимулы приводят к более сложной деятельности мозга и постепенному увеличению мозга.
- Изменения в конструкции зубов и зубных дугах.
- Резаки и клыки сжимаются и не отделяются от стульев.
Клиновидная кость (os sphenoidale) лежит в основании мозгового черепа и соединяется со всеми его костями (Атл., 16, А), как бы вклиниваясь между ними, но по форме она с клином не сходна. Кость имеет сложное строение, так как через нее проходит много крупных нервов, она участвует в образовании глазницы, височной и подвисочной ям, служит местом прикрепления жевательных мышц.
Однако некоторые особенности специализации могут наблюдаться в отдельных строках. Важной особенностью приматов является их адаптация жизни на деревьях. Поэтому признаки жизни в этой среде развились, такие как очень быстрые реакции на внешние раздражители и бинокулярное зрение. Напротив, было сильное подавление запаха.
Все адаптивные символы отражены в скелете приматов. Череп имеет большие размеры, особенно его череп. Глаза большие и не расположены по бокам, а вперед. Остальные части черепа разделены костной стенкой. Из-за плохого запаха нос обычно имеет небольшие размеры. Челюсти короткие, а зубы имеют примитивную структуру. Передние и задние конечности очень подвижны.
В кости различают тело с воздухоносной пазухой, которая сообщается спереди с носовой полостью (Атл., 15, Б). Углубление на верхней поверхности тела называют турецким седлом , ограниченным сзади спинкой седла; в нем помещается железа внутренней секреции - гипофиз. В обе стороны от тела отходят большие крылья ; в основании каждого из них последовательно расположены круглое, овальное и остистое отверстия . Передняя поверхность крыльев образует латеральную стенку глазницы. Выше больших крыльев от тела кости отходят малые крылья , пронизанные у основания зрительным каналом , в котором расположен одноименный черепной нерв. Малые крылья отделены от больших верхнеглазничной щелью и участвуют в образовании глазницы. Вниз от тела отходят крыловидные отростки , состоящие из двух (медиальной и латеральной) пластинок, между которыми находится крыловидная ямка . Основание отростков пронизано крыловидным каналом . Отростки служат местом прикрепления мышц.
Анатомические особенности также следует распространить на развитие потомства после рождения. Это расширяет контакты между матерью и матерью, которая играет большую роль в обучении и передаче знаний. Даже черепа самых старых полупапов отличаются от архаичных млекопитающих. Они показывают большое боковое расширение мозга в области слуховой доли и отчетливое развитие маленького мозга. Ископаемые результаты палеогена указывают на относительное увеличение отдельных частей мозга. Увеличение произошло особенно в области сна, лобная область остается относительно небольшой.
Решетчатая кость (os ethmoidale) окружена другими костями так, что на целом черепе видна лишь ее наружная часть - глазничная пластинка , участвующая в образовании медиальной стенки глазницы (Атл., 16, Б). Другая часть кости - решетчатая пластинка - замыкает вырезку лобной кости и видна с мозговой поверхности черепа. От этой пластинки вверх отходит продольный петуший гребень ; продолжением его в носовую полость служит перпендикулярная пластинка , участвующая в образовании перегородки носа (Атл., 15, Б). Большие парные части кости - лабиринты , состоящие из костных воздухоносных ячеек, свисают в носовую полость. В сторону перпендикулярной пластинки от лабиринтов выступают средняя и верхняя носовые раковины .
Увеличение этих участков произошло позже. Можно спросить, связан ли рост мозга млекопитающих с некоторыми явлениями развития или был ли он спонтанным. Исследования показали, что увеличение мозга происходит в половине ушей в самом начале их излучения, а затем в среднем эоцене. Можно утверждать, что оба явления коррелированы и поэтому зависят друг от друга. Увеличение мозга во время третичного периода после длительного застоя во время мезозоя характерно для всех млекопитающих. Мозг птиц, например, рос намного быстрее, чем мозг млекопитающих.
Лицевой отдел . В лицевом отделе, в отличие от мозгового, преобладают парные кости, к которым относятся верхнечелюстные, носовые, слезные, скуловые, нёбные и нижние носовые раковины. Непарных костей всего три: сошник, нижняя челюсть и подъязычная кость (Атл., 14 и 15).
Верхнечелюстные кости (maxilla) самые большие парные кости в лицевом черепе, занимающие центральное место в лицевом черепе; они состоят из тела и четырех отростков (Атл., 17). Внутри тела находится большая воздухоносная верхнечелюстная пазуха , открывающаяся в носовую полость. Передняя, лицевая поверхность тела вогнута, имеет на себе кыковую ямку , а над ней - нижнеглазничное отверстие одноименного канала, пронизывающего всю кость. Верхняя поверхность тела образует нижнюю стенку глазницы, а носовая поверхность - боковую стенку носовой полости. К этой стенке прикрепляется небольшая кость - нижняя носовая раковина . Задняя поверхность кости обращена к подвисочной яме. Из четырех отростков , отходящих от тела, лобный соединяется с лобной, а скуловой - со скуловой костью. Нёбные отростки вместе с прилегающими к ним сзади нёбными костями (ossa palatina) образуют твердое нёбо . Альвеолярный отросток снабжен восемью лунками, в которых сидят верхние зубы.
Увеличение мозгового млекопитающего относительно быстрое во время триаса при переходе от уровня рептилии к уровню млекопитающих. Около 120 миллионов лет в мезозое мозг млекопитающих почти не рос. У мезозойских и кенозойских млекопитающих были относительно небольшие мозги. Не исключено, что это было связано с адаптацией к стабильной среде, в которой не было никакого давления выбора, которое могло бы привести к росту мозга. В начале кенозоя и, возможно, в конце верхнего мела расширение мозга произошло только у приматов.
Еще до начала эоцена относительный размер мозга приматов сравним со средним размером мозга других млекопитающих. На основании ряда исследований можно показать, что эволюция мозга человека является биологическим явлением того же типа, как эволюция мозга всех других млекопитающих. В настоящее время количество видов грызунов превышает количество видов всех других млекопитающих. Существует около 350 видов грызунов и вымерших видов.
Носовые кости (ossa nasalia) расположены в области переносицы и замыкают сверху грушевидное отверстие , ведущее в носовую полость. В глубине последней виден сошник (vomer) сагиттально расположенная пластинка, прирастающая к ьлино видной, решетчатой, нёбным и верхнечелюстным костям.
Слёзные кости (ossa lacrymalia) - самые маленькие из костей лицевого черепа. Образуя часть внутренней стенки глазницы, они примыкают к лобной, решетчатой и верхнечелюстной костям.
Вероятно, многие виды грызунов также существовали в третичном состоянии. Несколько факторов способствовали успешному развитию грызунов. Грызуны оставались маленькими млекопитающими на протяжении всего их развития, хотя общая тенденция привела к постепенному увеличению размеров у всех других животных. Небольшой размер был благоприятной адаптацией к окружающей среде, которая недоступна для крупных млекопитающих. По этой причине грызуны, похоже, очень быстро размножаются и создают большие популяции.
Мы находим грызунов на поверхности земли, под землей, на деревьях, на скалах, в болотах, на лугах и на всей планете. Образ жизни привел к развитию двух пар резцов в верхней и нижней челюстях. Эти зубы растут на протяжении всей жизни животного. Третий резчик и собака, общие для других млекопитающих, подавляются у грызунов. Большинство грызунов питаются растениями, некоторые едят насекомых или всеядны. У некоторых грызунов есть тело, которое позволяет вам прыгать.
Скуловые кости (ossa zygomatica) имеют по три отростка лобный, височный и верхнечелюстной , названные по костям, с которыми они соединяются. Скуловые кости образуют нижненаружные края глазниц, а вместе со скуловыми отростками височных костей - скуловые дуги .
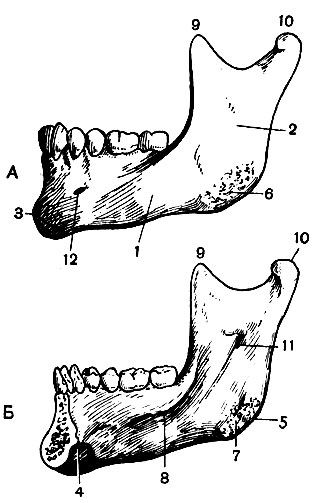
Нижняя челюсть (mandibula) - непарная кость, она состоит из тела и двух ветвей (рис. 44). Спереди на теле выдается подбородочный выступ , а по бокам его - подбородочные бугорки . На внутренней поверхности тела по средней линии находится подбородочная ость , от которой в стороны тянутся две черепно-подъязычные линии . На верхнем альвеолярном крае тела находится 16 зубных лунок. Отходящие от тела ветви образуют с ним угол , на внутренней и наружной поверхностях которого находятся шероховатости - места прикрепления жевательных мышц. Ветви заканчиваются двумя отростками; из них передний - венечный - служит местом прикрепления одной из жевательных мышц, а задний - мыщелковый (в котором различают головку и шейку) - сочленяется с височной костью. На внутренней поверхности ветви находится отверстие нижнечелюстного канала . Пройдя вдоль корней зубов, канал открывается на наружной поверхности тела подбородочным отверстием .
Подъязычная кость (os hyoideum) - маленькая изогнутая кость, подвешенная к шиловидному отростку височной кости при помощи длинной связки. Состоит из тела, малых и больших рожков (рис. 45). Эту кость легко прощупать на шее над гортанью.
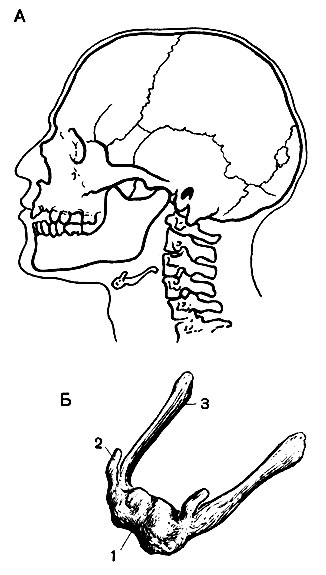
Соединение костей черепа . Почти все кости черепа соединены неподвижно швами . Это разновидность синдесмоза, где прослойка волокнистой соединительной ткани, связывающая смежные кости, едва заметна (Атл., 14 и 15). У взрослых, особенно у стариков, большинство швов окостеневает, но между пирамидой и соседними с нею костями сохраняется синхондроз. Единственная подвижно соединенная кость черепа (кроме подъязычной) - нижняя челюсть.
Височно-нижнечелюстной сустав (articulatio temporomandibularis) парный, комбинированный, образуется эллипсоидной формы головкой нижней челюсти, сочленовными ямкой и бугорком височной кости. Между сочленяющимися костями расположен двояковогнутый хрящевой диск; он разделяет суставную полость на верхний и нижний отделы. При движении челюсти вперед ее головки вместе с дисками передвигаются на сочленовные бугорки. В суставе возможны опускание и поднимание челюсти (вокруг фронтальной оси), смещение ее в стороны (вокруг вертикальной оси), что важно при жевании, а также движениях, связанных с членораздельной речью.
С позвоночником череп соединен атланто-затылочным суставом .
Топография черепа . Емкость мозгового отдела составляет у мужчин в среднем 1450 см3, у женщин она меньше и в среднем равна 1300 см 3 . Эта емкость связана с общими размерами тела.
Мозговой череп делится на крышу и основание (Атл., 14 и 15). Крыша образуется теменными костями, чешуей лобной, затылочной и височных костей, частью больших крыльев клиновидной кости. Остальные части этих костей и решетчатая кость формируют основание . Кости крыши плоские. На их вогнутой мозговой поверхности видны артериальные борозды, непостоянные мелкие отверстия, через которые проходят вены, следы широких венозных синусов и ряд уплощенных вдавлений и выступов, повторяющих рельеф мозга. Мозговая поверхность основания черепа также повторяет рельеф нижней поверхности мозга. Здесь различают переднюю, среднюю и заднюю черепные ямы (Атл., 15, А).
Передняя черепная яма (fossa cranii anterior) образована лобной костью и решетчатой пластинкой решетчатой кости; задней границей ее служат края малых крыльев и тело клиновидной кости.
Средняя черепная яма (fossa cranii media) отделяется от задней краем пирамиды височной кости и спинкой турецкого седла; она образована большими крыльями клиновидной кости, передней поверхностью пирамид и чешуей височной кости. Нижнебоковые части ямы пронизаны большим числом отверстий. Впереди турецкого седла расположены зрительные каналы, сбоку от них - верхнеглазничные щели (те и другие ведут в глазницу), а позади них - круглые отверстия, далее - овальные, а за ними - маленькие остистые отверстия. Между задним краем клиновидной кости (сбоку от ее тела) и пирамидой видно рваное отверстие , в области которого находится внутреннее отверстие канала сонной артерии.
Задняя черепная яма (fossa cranii posterior) почти целиком образована затылочной костью; лишь незначительные переднебоковые части ямы образуются задней поверхностью пирамид с сосцевидной частью височных костей. Центр ямы занимает большое затылочное отверстие , посредством которого полость черепа сообщается с позвоночным каналом. Латеральнее отверстия проходит канал подъязычного нерва, а впереди находится скат, образованный слиянием тел клиновидной и затылочной костей. На скате расположен продолговатый мозг. Между затылочной костью и пирамидой образуется яремное отверстие , спереди от него, на задней поверхности пирамиды, открывается внутреннее слуховое отверстие.
На наружной поверхности черепа различают две глазницы, вход в носовую полость, парные височные и подвисочные ямы.
Глазницы (orbitae) - полости, ограниченные четырьмя стенками. Их верхняя стенка образована лобной костью и малыми крыльями клиновидной; медиальная - слезной и решетчатой костями; нижняя - верхнечелюстной и отчасти скуловой костью, а латеральная - большими крыльями клиновидной кости и отчасти лобной и скуловой костями. Нижняя стенка отделена от латеральной нижнеглазничной щелью , через которую глазница сообщается с подвисочной ямой и крыло-нёбной ямкой. В глубине глазницы открываются верхнеглазничная щель и зрительный канал, ведущие в полость мозгового черепа. На стыке носового отростка верхнечелюстной кости со слезной начинается слезно-носовой канал , ведущий в носовую полость.
Носовая полость (cavum nasi) открывается спереди грушевидным отверстием , а сзади - двумя хоанами . В глубине ее хорошо видна костная носовая перегородка , которая состоит из сошника и перпендикулярной пластинки решетчатой кости. Полость снизу образована верхнечелюстными и нёбными костями, с боков, кроме того, еще слезными и решетчатой костями, крыловидными отростками клиновидной кости, а сверху - носовыми, лобной и решетчатой костями, телом клиновидной кости. В полость носа вдаются три носовые раковины (верхняя и средняя - выросты лабиринтов решетчатой кости, нижняя - самостоятельная), под которыми образуются верхний, средний и нижний носовые ходы . В верхний открывается пазуха клиновидной кости, в средний - лобная и верхнечелюстная пазухи, в нижний - слезно-носовой канал. Кроме того, в средний и верхний ходы открываются ячейки лабиринта решетчатой кости. Нёбно-клиновидным отверстием у заднего конца верхней раковины полость носа сообщается с крыло-нёбной ямкой.
Височная яма (fossa temporalis) образована боковыми частями черепа и ограничена снаружи скуловой дугой, ниже которой переходит в подвисочную яму.
Подвисочная яма снаружи прикрыта ветвью нижней челюсти, сообщается со средней черепной ямой через овальное и остистое отверстия. В глубине подвисочной ямы открывается воронкообразная крыло-нёбная ямка . В последнюю из средней черепной ямы ведет круглое отверстие, из носовой полости - нёбно-клиновидное, из глазницы - нижнеглазничная щель. Книзу крыло-нёбная ямка переходит в узкий крыло-нёбный канал , открывающийся на твердом нёбе.
В местах, где череп испытывает давление при жевании, возникают приспособления в виде плавно изогнутых утолщений компактного вещества, выступающих над поверхностью кости, - контрфорсы . Особенно важны два контрфорса - носо-лобный и скуловой . Оба упираются в альвеолярную часть верхнечелюстной кости, причем первый поднимается от клыка через ее лобный отросток к носовой части лобной кости, а второй идет от коренных зубов через скуловую кость на скуловые отростки лобной и височной костей. Контрфорсы обеспечивают передачу давления нижней челюсти на верхнюю и равномерное распределение его на мозговой отдел черепа.
Пазухи , залегающие в верхнечелюстных височных, лобной и клиновидной костях, увеличивают объем черепа и в то же время значительно облегчают его без особого изменения крепости и других механических свойств, а также служат резонаторами звуков речи. У новорожденных пазухи выражены слабо. Например, наиболее емкая из них - верхнечелюстная - представляет у младенца лишь небольшое впячивание боковой стенки носовой полости. Постепенно развиваясь, пазухи достигают своей постоянной величины лишь с окончанием роста черепа. Все пазухи содержат воздух, поступающий в них из носовой полости (кроме ячеек сосцевидного отростка), и называются воздухоносными , а кости, содержащие их, - пневматизированными .
Онтогенез черепа . В черепе человека одни кости возникают путем замещения хряща, а другие - как покровные. Образование покровных костей происходит уже с середины второго месяца утробной жизни, тогда как замещение хряща костью начинается позднее - с конца этого месяца или даже со следующего. Все очаги окостенения возникают в определенном порядке, и сначала число их велико, например в клиновидной кости достигает десяти. Слияние очагов происходит уже в утробной жизни и продолжается после рождения.
Покровные кости крыши черепа плода, разрастаясь, сближаются друг с другом, но и у новорожденного между ними остаются еще перепончатые соединительнотканные участки. Такие участки особенно велики там, где сходится несколько покровных костей, и называются родничками. Роднички располагаются по углам обеих теменных костей, вследствие этого образуются непарные лобный и затылочный и парные передние боковые и задние боковые роднички (рис. 46). Благодаря такому соединению кости крыши черепа могут заходить своими краями друг на друга. Поэтому череп зрелого плода способен изменять свою форму и приспосабливаться к диаметру малого таза при прохождении через родовые пути. У детей затылочный родничок зарастает на втором месяце жизни; наибольший, лобный легко прощупывается и у нормально развивающегося ребенка полностью зарастает лишь к полутора годам. Помимо родничков, у новорожденных между некоторыми костями основания черепа сохраняется и хрящ.
У новорожденного череп относительно (к размерам всего тела) намного больше, чем у взрослых. Голова у него - наибольшая по периметру часть тела и именно поэтому даже при нормальном положении плода труднее всего проходит через родовые пути. Высота головы (от темени до подбородка) у новорожденного укладывается в длине тела четыре раза. С возрастом это соотношение меняется, так как рост черепа (головы) отстает от роста всего организма. У взрослого высота головы составляет лишь 1 / 8 длины тела.
Новорожденный не имеет прорезавшихся зубов, поэтому челюсти его развиты слабо. Мозговой череп вследствие энергичного развития мозга и органов чувств относительно велик и резко преобладает над лицевым (5:1). Его емкость 385-450 см 3 . Кости тонки, в своде гладки и гибки, теменные бугры выступают резко, но места прикрепления мышц и связок не рельефны, а сосцевидный отросток височной кости едва заметен (как у антропоидов). Нижнечелюстная кость состоит из двух половин (рис. 47, 1), срастающихся в течение второго года жизни. Лобная кость также состоит из двух частей (полностью сливающихся лишь к шести годам), затылочная - из пяти, а височная - из четырех (рис. 46).
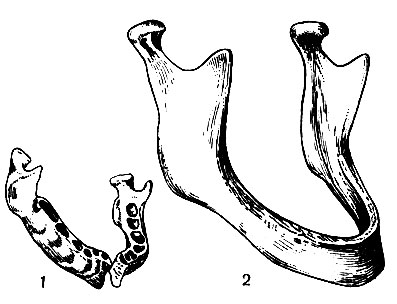
Возрастные изменения черепа у детей заключаются в росте и слиянии отдельных костных очагов. После прорезывания зубов лицевой отдел начинает расти быстрее мозгового, что ведет к заметным изменениям в пропорциях частей черепа. Хотя рост черепа заканчивается к 23-25 годам, изменение его продолжается до старости. После 30 лет швы начинают зарастать. В старческом возрасте кости становятся более тонкими, легкими и губчатое вещество их разрыхляется, резорбируется. В связи с выпадением зубов рассасываются зубные края челюстей (рис. 47, 2), высота лицевого отдела значительно уменьшается, и над ним опять начинает резко преобладать мозговой череп.
Формы черепа . Индивидуальные колебания формы черепа довольно значительны. В частности, по внешней форме выделяют черепа узкие, длинные (долихокрания), среднекруглые (мезокрания) и круглые, широкие (брахикрания).
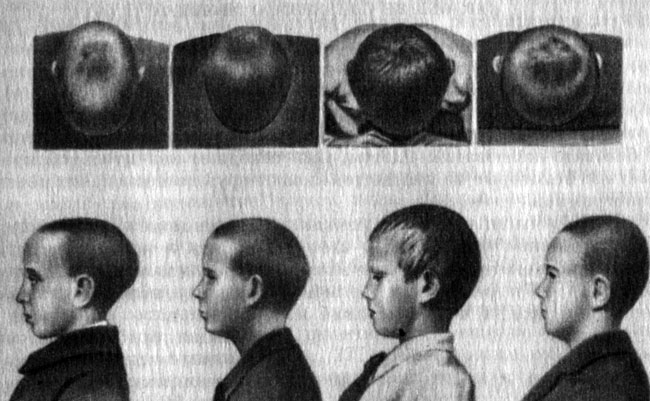
В прошлом некоторые народы искусственно деформировали головы у детей тугим бинтованием, вследствие чего череп постепенно приобретал причудливую форму - конусовидную, башенную и т. п. (рис. 49).
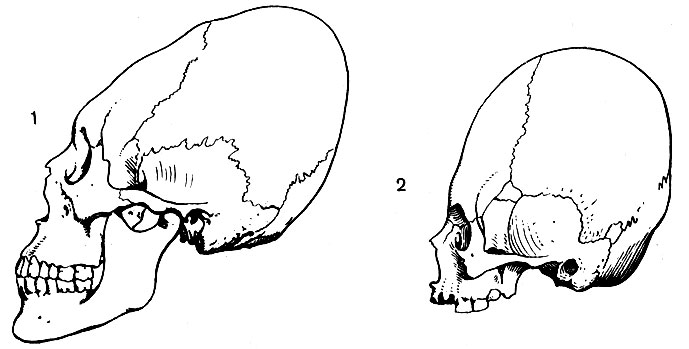
Отличить череп мужской от женского не всегда удается, так как половые различия в его форме и строении часто выражены нерезко. Череп мужчины в среднем несколько больше, на нем сильнее выражены места прикрепления мышц и надбровные дуги.
Скелет млекопитающих отличается большой прочностью и разнообразием строения у разных видов в связи с условиями их обитания и способами передвижения. Упрочение скелета достигается срастанием костей , особенно в черепе, увеличением массы костной ткани и усложнением её структуры .
Трубчатые кости , несущие основную нагрузку, характеризуются слоистым строением. Их стенки сложены из так называемых остеонов, между которыми располагаются вставочные костные пластинки. Остеоны представляют собой вложенные друг в друга трубочки, состоящие из пластинок, соединённых пучками коллагеновых фибрилл. Внутри каждого остеона имеется полость – гаверсовый канал , в котором проходят нерв и кровеносный сосуд, обеспечивающий доставку питательных веществ и кислорода. Снаружи кости одеты плотной соединительнотканной оболочкой, которая носит название надкостница , или периост. Через неё проходят нервы и кровеносные сосуды к гаверсовым каналам костей.
Внутри костей расположен костный мозг , являющийся органом кроветворения. Концы трубчатых костей (эпифизы) состоят из губчатой ткани. Прочность костей во многом определяется степенью их минерализации – содержанием кальциевых солей в костной ткани. Она наиболее велика у наземных видов, особенно обладающих большой массой в сочетании с быстрым передвижением.
Позвонки, слагающие осевой скелет, имеют плоские сочленовные поверхности с хрящевыми прослойками (платицельные).
Череп
Череп млекопитающих характеризуется большим объёмом мозговой коробки, двумя затылочными мыщелками, уменьшением числа костей за счёт их редукции и срастания. Череп платибазальный (с широким основанием и расставленными глазницами, между которыми находится мозговая полость). На наружной части костей, особенно в затылочной области, развиты гребни (выступы костей), предназначенные для прикрепления мускулатуры.
Затылочный отдел мозгового черепа состоит из одной затылочной кости, образованной срастанием верхней затылочной, основной и двух боковых затылочных костей (рис. 79). В месте их соединения формируется большое затылочное отверстие, служащее соединению головного и спинного мозга. В нижней части затылочного отдела расположены два мыщелка для прикрепления черепа к позвоночному столбу.
Крыша черепа состоит из парных носовых, лобных, теменных костей и непарной межтеменной кости, которая у многих видов прочно срастается с примыкающими к ней костями.
Бока черепа формируют: крупная чешуйчатая кость, каменистая кость (образована срастанием ушных костей), костные элементы глазницы (глазо-клиновидная, крылоклиновидная, слёзная) и скуловая дуга , ограничивающая глазницу снизу.
В отличие от диапсидного черепа рептилий (имеется две скуловые дуги), череп млекопитающих относится к синапсидному типу (одна скуловая дуга, но смешанного типа). Она сформирована соединением скуловой кости с верхней челюстью и скуловым отростком чешуйчатой кости. Область среднего уха прикрыта барабанной костью (слуховой барабан), возникшей эволюционно из угловой кости рептилий. Срастание чешуйчатой, каменистой и барабанной костей образуют общую височную кость .
Дно черепа образовано парными нёбными, крыловидными костями и непарным сошником. В укреплении дна черепа принимают участие и кости нижней части орбиты глаза (основная и передняя клиновидные ), кпереди от которых лежат решетчатая кость (обонятельная) и носовая перегородка .
Для всех зверей свойственно твёрдое костное нёбо , сформированное срастанием нёбных костей с отростками верхнечелюстных и межчелюстных костей. В задней его части имеется соединительнотканная перепонка, или мягкое нёбо , за которым расположены хоаны. Вторичное нёбо позволяет животным беспрепятственно дышать при приёме пищи.
Висцеральный череп млекопитающих также претерпел значительные изменения. Верхние челюсти , состоящие из сросшихся межчелюстных и верхнечелюстных костей, глубоко врастают в мозговой череп. Нижние челюсти образованы зубными костями, которые имеют венечные, сочленовные и угловые отростки . Сочленовные отростки зубных костей соединяются со скуловыми отростками чешуйчатых костей и, таким образом, подвешивают нижние челюсти непосредственно к мозговому черепу.
Квадратные и сочленовные кости , выполнявшие у низкоорганизованных позвоночных функцию сочленения челюстного аппарата с мозговым черепом, у млекопитающих преобразованы в слуховые косточки среднего уха . Из квадратной кости сформирована наковальня , а из сочленовной кости – молоточек. Верхняя и нижняя челюсти имеют особые углубления (альвеолы), в которых сидят зубы.
Рис.79. Череп лисицы. А – вид сбоку; Б – вид снизу:
1 – затылочная кость, 2 – большое затылочное отверстие, 3 – затылочный мыщелок, 4 – межтеменная кость, 5 – теменная кость, 6 – лобная кость, 7 – надглазничный отросток лобной кости, 8 – носовая кость, 9 – височная кость, 10 – скуловой отросток височной кости, 11 – скуловая кость, 12 – основная клиновидная кость, 13 – передняя клиновидная кость, 14 – сошник, 15 – предчелюстная кость, 16 – небный отросток предчелюстной кости, 17 – верхнечелюстная кость, 18 – небный отросток верхнечелюстной кости, 19 – небная кость, 20 – крыловидная кость, 21 – слезная кость, 22 – барабанная кость, 23 – зубная кость, 24 – венечный отросток зубной кости, 25 – резцы, 26 – клык, 27 – предкоренные зубы, 28 – коренные зубы, 29 – наружный слуховой проход
Подъязычный аппарат представлен подъязычной костью с передними и задними рожками.