Szag. Az orrüregben szaglás szervek vannak.
Isolation. Néhány anyag (hulladéktermékek stb.) A légzőrendszeren keresztül szabadulhat fel.
Védő. Jelentős számú specifikus és nem-specifikus immunformáció létezik.
A hemodinamika szabályozása. Belélegezve a tüdő növeli a vénás vér áramlását a szívbe.
Sajnos az utolsó kérdéssel kapcsolatos bizonyítékok nem könnyűek. Mindazonáltal az értelmezés bizonyos következménye a madár hangosításával kapcsolatos. A madarak, akárcsak az emberek, énekhangját olyan készülék vezérli, amely nem igényli független mozgást a bal és a jobb oldalon.
Lehet, hogy a beszédkészülék két felének egyenlő, de független megjelenítésének hiánya fontos tényező az emberi agy aszimmetriájának kialakulásában, mivel hasonló aszimmetriák fordulnak elő a dalmadarakban. A vizsgált madarakban a hangképző szervnek, a syrinxnek a bal és a jobb oldalán izmok vannak, és mindegyik készlet a hipoglobularis koponya ideg különálló ágára reagál. A fecskendő izmainak bal oldali halmaza sokkal nagyobb, mint a jobb, ami önmagában is sugalmazó, de egyértelmű eredményeket érhetünk el, ha elválasztjuk az ideget a bal izomtól, és összehasonlítjuk ennek a műveletnek a hatásait, ha az ideget a fecskendő jobb oldalán lévő izmokra vágjuk.
Depot vér.
A hőszabályozás.
Halak, kétéltűek, hüllők, madarak, emlősök és emberek légzőrendszerének jellemzői.
A ciklosztómák, porcok, csonthalak légzőrendszerének jellemzői
Más gerinces állatoktól eltérően, a kopolytómák a kopoltyúrésekben endodermális eredetű kopoltyúkat fejlesztenek ki. A kopoltyúzsákok belső felülete a nyálkahártya számos redőjét képezi, amelyben a kicsi erek sűrű hálózata van - a kapillárisok. A máglyában minden egyes ág (összesen 7 pár) kifelé nyílik, független külső nyílással.
Ha a metszés bal oldalon van, akkor a kanári által használt korábban használt összes szótag vagy csaknem minden szótag elveszik, míg a jobb oldal izmainak inaktiválása esetén az éneklés nem változik szinte azonnal utána, csak egy vagy két szótag hiányzik a legtöbb.
Mivel a kanári bal oldali hypoglossális idege az agy bal oldaláról származik, feltételezhető, hogy a bal oldali agyban a vokalizáció domináns a kanáriban, és más kísérletek megerősítik, hogy ez így van. Az embernek van egy bal oldali félgömb homloklebenyének területe, amelyet Broca területének neveznek, mert egy ilyen nevű francia neurológus megállapította, hogy a sérült betegek afázia formájában szenvedtek. A külső striatumban található, amelyet más okokból hasonlónak tekintünk az emlősök agykéregében.
A kopoltyúzsák belső nyílása összekapcsolja üregét a légzőcsővel, amely egy vak kinövék, elõtt a szájüreghez csatlakozik, és mozgatható redõvel - vitorla határolja. A lebegő lámpánál a víz belép a szájüregbe, ahonnan átjut a légzőcsőbe, és a kopoltyúzsákokon áthaladva a külső nyílásaikon keresztül kiszökik. Ebben az esetben a vízben feloldott oxigén behatol a kapillárisokba, és megkötve van a vérpigmenttel, és a vénás vért telítendő szén-dioxid átjut a vízbe, és azzal együtt kivezetik.
Noha a bal oldali féltekén ezen a területen a sérülések csaknem az egész dal azonnali veszteségéhez vezettek, a jobb féltekének megfelelő pontján elhelyezett sérülések lehetővé tették a szokásos szótagok több mint felének azonnali reprodukcióját közvetlenül a műtét után olyan formában, hogy az emberi fül megkülönböztethetetlen a műtét előtt. Az emberekkel szenvedő agykárosodásból eredő nyelvi károsodás előrejelzése nem jó, de a spontán módon vagy a logopédiai kezelés során sem ritka a jelentős gyógyulás, különösen viszonylag tiszta balesetek vagy katonai sérülések, nem sokk helyett. vagy daganatok.
Abban az esetben, ha a májat etetik (vagy bármilyen tárgyhoz ragasztják), a szájüregben a víz áramlása lehetetlenné válik. Ugyanakkor a vitorla víz áthaladása nélkül bezárja a légzőcső bejáratát, és a folyékony étel a szájüregből átjut a nyelőcsőbe. Ebben az esetben a légzést más módon hajtják végre: a testfalak izmainak hatására a kopoltyúzsákok összenyomódnak, és a vizet a külső kopoltyúnyílásokon keresztül tolják (aktív kilégzés); ezenkívül a porcos kopoltyúrács rugalmassága miatt a kopoltyúrégió ismét tágul, és ugyanazon külső lyukakon keresztül a vizet ismét szívja a kopoltyúzsákokba (passzív belélegzés).
Az emberi nyelv mechanizmusaival egy másik párhuzamos az, hogy ha a hypinlobularis ideg bal oldali ágát, amely a syrinxbe megy, fiatal madarak vágják le, akkor a jobb struktúrák nagyon gyorsan átveszik az irányítást a dalban. Függetlenül attól, hogy a hangok hallható megértése vagy megértése ugyanolyan oldalirányú-e egy dal-madár agyában, mint amilyennek látszik, a vokális produkció továbbra is fennáll. Egy teljesen más faj akusztikus lateralizációja több bagolyfajban ismert, amelyek közül néhánynak a külső fül fizikai aszimmetriája nagyon egységes, minden egyénnél.
A porcos halak légzőrendszerének fő részét ektodermális eredetű nagy kopoltyúlemezek alkotják, amelyek az egyik oldalukon vannak rögzítve a kopoltyú válaszfalaihoz. Belélegezve a garat kiszélesedik, a szájon keresztül víz jut és mosja a kopoltyúlemezeket. Kilégzéskor a garat térfogata csökken, és a kopoltyúhoronykon keresztül a víz kijön, ezek közül 5-7 van. A kartigénes halaknak nincs kopoltyúfedele. A kopoltyúrésen kívül a szem hátsó nézetének legtöbbször van olyan kezdetleges kopoltyúrés - fröccsent, amely a garat elülső oldalán nyílik, ahol belélegezve a víz behatol rajtuk keresztül.
Kíváncsi lenne, ha az összes gerinces ember közül csak dalmadarak, baglyok és emberek használnák ki a kétoldalú agyszimmetria relaxációját. Sajnos még mindig nem egyértelmű, hogy az emberi agy oldalsó funkciója más főemlősök előidézője-e, és ez nyilvánvalóan döntő pillanat. Az anatómiai egyenlőtlenségeket illetően Cunningham felfedezte, hogy az emberi zsíros hasadások aszimmetriái vannak a nagy majmok és a nagy majmok közötti hasonlóságokban, és ezt Yeni-Comanshan, Benson, Lemey és Geschwind, Kain és Wada igazolta.
Véleményem szerint jelenleg a legjobb becslés az, hogy az agyműködés aszimmetriája önmagában nem egy személy előjoga, bár az oldalirányú dolgok egyike a nyelv. Az agyi régió laterálissága sem szükséges, sem nem elégséges feltétel.
A csontos halak légzőrendszerének fő szervei kopoltyúk, amelyek számos sziromból állnak, kopoltyú ívekhez közeli végükön, ellentétben a porcos halak kopoltyúlapjaival, amelyek egyik oldalán a kopoltyú válaszfalai vannak rögzítve. Következésképpen a csonthalak kopoltyúinak felülete sokkal nagyobb, mint a porcos halak. A kopoltyúk mozgatható fedéllel fedik le és védik.
Az emberi szellemi tevékenység szempontjából, ezért nem szabad azt feltételezni, hogy az agy oldalsó szintje határozza meg az állati faj ismeretének mértékét. Hierarchikus tervezés gerincesekben. A filogenetikai elmélet, az emberi neurológia és az agy működésének általános leírását célzó más kísérletekkel spekulatív egyszerűsítés az agyi aktivitási szintek hierarchiájának azonosítása. A legalacsonyabb szinten a gerincvelő egyértelműen biztosítja a közvetlen útvonalat az érzékszervi bemenet és a motor kimenete között.
Belégzéskor a szájüreg kibővül, és a benne lévő nyomás csökken, amelynek eredményeként a víz belép a szájnyíláson. Ugyanakkor a víz nyomása alatt a kopoltyú fedele hátsó vége bezárul, megakadályozva a víz áramlását ezen az oldalon. Ugyanakkor a kopoltyúfedél izmai összehúzódnak és a kopoltyú ürege tágul. Kilégzéskor a szájnyílás és a nyelőcső bemeneti nyílása bezáródik, és a szájüreg alja megemelkedik. Ennek eredményeként a vizet a kopoltyúrésen át átnyomják a kopolyaüregbe, és a kopoltyúk feletti áthaladása után kifelé megy a kopoltyúfedél hátsó széle közelében, amely most víznyomás alatt nyitva van.
A legmagasabb szinten az emlőskéreg bizonyos területei érzékszervi információkat kapnak, amelyeket később átalakítanak, kiszűrnek és osztályoznak az előző szakaszokban, míg a kéreg más területei olyan intézkedéseket kezdeményeznek, amelyeket az agy alsó részeinek részletesebb szervezése adhat.
A gerincvelő és az agykéreg, agy középső agya, a kisagy, a thalamus és a test striatuma között specifikus közvetítő szerepek rendelhetők. Nyilvánvaló, hogy az alapfeltevés az, hogy az agyi szervezettség magasabb szintje a megismerés és a gondolkodás komplex formáival jár, míg az alacsonyabb szint csak a mechanikusabb és reflektívabb kérdésekre vonatkozik. Először, sok fontos részletet szükségszerűen figyelmen kívül hagyunk: a funkciószintek meghatározása megkönnyíti a dolgok megértését, de a szintek bizonyos szempontból kényelmesek lehetnek.
A légzőrendszer szövődményei
Szinte minden kétéltűnek tüdeje van. Mivel a tüdő belső fala szinte sima, felületük viszonylag kicsi. A légcső szinte fejletlen, a tüdő közvetlenül kapcsolódik a gégéhez. Mivel a kétéltűeknek nincs mellkasuk, a légzést a szájüreg izmai biztosítják. Nyitott orrlyukakkal (amelyek - a hal-orrlyukaktól eltérően - nyitottak, vagyis a külső orrlyukakon kívül vannak még belső orrlyukak is) és a zárt szájjal a hatalmas szájüreg alját meghúzzák, és a levegő belép. Ezután az orrlyukakat speciális szelepekkel lezárják, a száj alja felemelkedik, és a levegő pumpálódik a tüdőbe. A kilégzés a hasi izmok összehúzódásának következménye. A kétéltűek jelentős mennyiségű oxigént kapnak a bőrön és a szájüreg nyálkahártyáin keresztül. Bizonyos típusú szalamandra egyáltalán nincs tüdeje, és a gázcsere a bőrön keresztül történik. A bőr azonban csak abban az esetben képes ellátni légzési funkciókat, ha nedves.
Másodszor, ha megosztják a funkciókat, akkor szinte elkerülhetetlen, hogy valaki engedelmeskedjen annak a kísértésnek, hogy hamis értékeket szabjon meg az osztásokra. Figyelmeztető példaként, az érzékszervi és a motoros funkciók elválasztását is néha gyanakvással tekintik. 4. ábra A gerinces agy részének elnevezése és általános terve. A felső ábra egy durva vázlat a gerinces agyban általában azonosított struktúrák közötti anatómiai viszonyokról. Az alsó ábra vázlatosan szemlélteti a fő szenzoros idegek belépési pontjait és az agy idegi kapcsolatainak jelentősen egyszerűsített útvonalait.
Ezért lehetetlen a kétéltűek élőhelye nem megfelelő páratartalom mellett. A vízben élő lárvák kopoltyúkkal (először külső, majd belsőleg) és a bőrrel lélegeznek. Néhány, a vízben állandóan élõ kétéltû kétéltûnek élettartama a kopoltyúk. Így a kétéltű légzési módszerek továbbra is közel állnak a halakhoz. A hüllők légútja a külső orrnyílásokkal - az orrlyukkal - kezdődik. Ezenkívül az orrcsatornán és a belső orrlyukakon áthaladó levegő - csontok - bejut a szájüregbe. A szájüreg mélyén a gége. Speciális izmokkal van felszerelve, és össze van kötve a hyoid készülékkel. A szájüregből a gégén keresztül belélegzett levegő bejut a légcsőbe.
Emlősökben és madarakban az előagy nagyobb, mint az itt megadott, és az agy egésze egy hosszú tengely mentén zsugorodik. Emlősökben a post-thalamikus vetületek az agyféltekének neocortexéből állnak. Az érzékszervi és motoros rendszerek összetettsége. Az agyszervezés értelmezésével az érzékszervi bemenetek szerint felfelé haladva a hátsó agy, a középső agy és az agy szintjén, fokozatosan távolabb távolabb magától az érzékektől, tervezett tevékenységekkel, összehangolt válasz-szekvenciákkal és a perifériás izmokhoz visszatérő mechanikus reflexekkel, az egyik azzal fenyeget, hogy az egyszerű input és output kategóriákat túl hangsúlyozzák.
A légcső a nyak mentén és a mellkasüregben fut, körülbelül a szív szintjén, és két rövid, a tüdőbe belépő hörgőkre oszlik. A tüdő vékonyfalú, üreges táska. A hüllők kétéltű tüdőjéhez képest komplexebb belső szerkezetük van: belső faluk, amelyben a kapillárisok elágaznak, szivacsos szerkezetűek, ami növeli a tüdő általános légzőfelületét. A hüllők egyetlen tüdője a tüdő. Ezen állatok bőre száraz, kanos mérleggel és keratinizált hámréteggel borítva, és nem vesz részt a légzésben. A gyíkok légzése a mellkas kiterjesztésével és összehúzódásával jár, speciális izmok hatására.
Ennek ellensúlyozására emlékeztetni kell arra, hogy az agy és a gerincvelő idegsejtjeinek 99% -a nem sorolható szenzoros vagy motoros állapotba, hanem „közbenső hálózatként” tekinthető a sejt szigorúan szenzoros vagy szigorúan idegrendszere között. Magától értetődik, hogy ami az érzékelő sejt axonok lágyításával kezdődik, és az izmok összehúzódásával végződik, az idegrendszerben hatalmas puffadásnak van kitéve, mielőtt ennek a folyamatnak a fordulása konvergenciává vált, és viselkedést eredményez.
Az érzés és az akció között a legprimitívesebb gerinces agyának is van saját élete. Az érzékszervi információk több célállomásra történő forkálását jól szemlélteti a vizuális útvonalak Ebbesson áttekintése. Ez azt mutatta, hogy a szálak kiindulási vetülete a szem retinajából öt vagy hat különböző pontra megy végbe a gerincesek összes osztályában. Ha nem vesszük figyelembe a többségüket, amelyek meglehetősen szűk funkciós sávokkal rendelkeznek, akkor még két fő vizuális útvonal létezik. Az egyik a szemtől a látótektómáig történő vetítéssel kezdődik a középső agyban, és általában úgy tekintik, hogy az alsó gerinceseket uralja, míg a másik a retina kivetítésével kezdődik a talamuszra.
A madarak légzőrendszerének jellemzői. A légzés mechanizmusa madarakban
Madarakban a levegő átjut az orrlyukakon és megcsavarodik a szájüregbe. Ezenkívül a légzőrendszer áthalad a gége hasadékán és a felső gégén, és tovább a légcsőbe. A madarak nyakának meghosszabbodása miatt a légcső sokkal hosszabb, mint a hüllőknél. A testüregbe belépve a szív szintjén két hörgőkre oszlik, a jobb és a bal tüdőbe áramlik, és ott erősen elágazik. A légcső alsó része és a hörgők kezdeti részei alsó gégét alkotják, amely csak a madarak számára jellemző vokális készülék. A madarak páros tüdeje kicsi és szorosan kapcsolódik a gerincoszlop oldalán lévő bordákhoz. A tüdő sűrű, szivacsos felépítésű.
Minden gerinces ember használja ezt a kettős látást, talán azért, mert az egyik út mozgó tárgyakat észlel, a másik pedig a rögzített mintákat elemzi. De mindegyik esetben a magban több mag is részt vesz, és rendkívül összetett átalakulások és absztrakciók történhetnek az agyféltekén, még az alsó gerinces állatokon is, mint például a cápák.
Legalább két párhuzamos, de egymást kiegészítő értelmezés használata az érzékenység bevitelére egy modalitás esetén a szabály, nem. Az egyes emlősök füléből származó akusztikus információk az agytörzs két magjára kerülnek, és tőlük vagy közvetlenül a talamuszba, vagy egy külön úton jutnak el a középső agyhoz, majd átjutnak a talamuszon a hallókéregbe. Hasonlóképpen, köztudott, hogy az emlősöknek a test felületének legalább két vetülete van a kéregben. A járművek mozgása kettős vagy hármas sávon is áthalad, mielőtt az izmokhoz konvergálnak, és ez a fő különbség a motorkéregből a gerincvelőig tartó piramis út és az extrapiramidális lánc között az előajj nem-kortikális struktúráin, valamint a középső agy és agy között, megkerülve. mindkettő vállalta, hogy felvegye a fejlõdési találkozókat a kisagyban.
A légzsákok elhagyják a madarak tüdejét. A madarak légzését a mellkas mozgatásával hajtják végre. A bordák izmainak összehúzódása és a bordák mozgékonysága miatt a szegycsont test elmozdul a gerincoszloptól, növekszik a test üregének térfogata, rugalmas légzsákok vannak nyújtva, és a levegő beszívódik a tüdőbe és a hátsó légzsákba (mellkas és hátsó rész); az első légzsákok (nyaki, interclavicularis, mellkasi mellkasi) nyújtásánál a tüdőből levegőt szívnak be. Kilégzéskor a szegycsont a gerincoszlop felé mozog, a szegycsont test és a belső szervek összenyomják a légzsákot, és a tüdőn már áthaladó levegőt az első légzsákokból kihúzzák a légcsőbe, ki, és a hátsó légzsákokból friss levegőt pumpálnak a tüdőbe. Tehát a madarakban szinte folyamatosan - mind az inspiráció, mind a kilégzés során - a tüdőn keresztül egy irányban friss levegő áramlik.
Nyilvánvaló, hogy a központi idegrendszer gazdagságának és sokszínűségének nagy része elveszik, ha a funkciót érzékelésre, cselekvésre és korrelációra kondenzáljuk: az „érzékszervi” és a „motor” kifejezéseket nem az alapkategóriák címkéjeként kell használni, hanem a problémák szinonimáiként. gyakorlati megoldások, amelyek megkövetelik egy komplex szerv, például az agy fejlődését, és amelynek értelmezése a neuropszichológiai elméletben még előzetes.
Az agy szervezetének filogenetikai változásainak hipotézisei. Figyelembe véve az alapfogalmak korlátozásait, most részletesebben beszélhetünk arról, hogy a gerincvelő agyának szervezete hogyan változhat osztályonként. Az encephalization doktrína leggyakoribb formája azt sugallja, hogy a viselkedéskezelési funkciók fokozatosan az agy magasabb szintjeire koncentrálódnak. Ezt a fogalmat alkalmazták a gerinces állatok gerinctelenekből történő evolúciójára, az evolúcióra.
A madarak légzőrendszerének ilyen jellegzetes felépítése a vér intenzív oxigéntelítettségét biztosítja. Az emlősökben a levegő a külső orrlyukakon keresztül jut a szaglási üregbe, onnan a chánon keresztül a garatba és a gégbe. A hangkábelek a gégében találhatók. A gége átjut a légcsőbe, egy hosszú csőbe, amely porcgyűrűkből áll, amelyek nem záródnak le a hátsó oldalon. A mellkasban a légcső két bronchusra oszlik, a tüdő felé tartva. A tüdőben a hörgők többször kisebb tubulusokra elágaznak; ezek közül a legkisebb vékonyfalú vezikulumokkal - alveolákkal - ér véget. A vérkapillárisok az alveolák falában vannak; éppen itt zajlik a gázcsere. A tüdő alveoláris felépítése csak az emlősökre jellemző.
Az egymást követő gerinces osztályok, valamint az emlősök rendjei és az azokon belüli különbségek. Emlősöknél a korticalizáció encephalizationnak tekinthető. Gyakran azt gondolják, hogy a főemlősök, fejlettebbek, szélesebb körben használják kéregüket, mint a rágcsálók vagy húsevők, és hogy egy másik főemlősök ebben a sorozatában az ember inkább a kortikális funkcióktól függ, mint Askim, majom vagy majom.
A hipotézis hipotézisének kialakulása az, hogy a központi idegrendszer bármelyik részét széles értelemben úgy lehet értelmezni, mint az érzékszervi információ értelmezését, valamint a viselkedés kontrollját és szervezését. Miután ezt megtették, egyre inkább válik, hogy valamilyen agyegységet jelöljön meg a többi „felelős” -ként, és ez előfeltételeket teremt a végrehajtó ág filogenetikai átvételéhez. Az agy bizonyos szintjének prevalenciája gyakran összefügg a korreláció, az integráció vagy az asszociáció kötelezettségével, tehát találhatunk hivatkozásokat a középső agyra, mint a korreláció domináns központjára, vagy a thalamusra, mint a legfejlettebb társulási mechanizmusra egy adott gerinces csoportban. A „domináns kontrollközpont migrációja” nyilvánvalóan a halak és az átlagos szárazföldi kétéltűek korrelációs és asszociatív tulajdonságainak seprésével jár a hüllő és a felső gerincvelő agya által.
A tüdő szabadon lóg a mellkasüregben lévő hörgőkön. Az egyes tüdőket lebenyre osztják, amelyek mennyisége az emlősök különböző fajtáinál változik. Az emlősök mellkasi üregét egyértelműen elválasztják a hasi üregtől egy folyamatos izomzattal - a membránnal. A légzést a mellkas és a membrán szinkron mozgatásával hajtjuk végre. Belégzéskor a mellkas ürege térfogata hirtelen növekszik a mellkas tágulása és a membrán ellapulása miatt; amíg a tüdő kinyúlik, szívva a levegőt. Kilégzéskor a mellkas falai egymáshoz közel helyezkednek el, és a membrán a kupola révén kinyúlik a mellkas üregébe. Ebben az esetben a mellkasüreg teljes térfogata csökken, növekszik a benne lévő nyomás és a tüdő összehúzódik, és a levegő kiürül belőlük.
Emberi és magasabb állatok légzési folyamatának jellemzői
A test energiaforrása a tápanyagok. A fő biokémiai reakció, amely felszabadítja ezen anyagok energiáját, az oxidáció, melyhez oxigénfogyasztás és szén-dioxid képződés társul. Az emberi testnek nincs oxigéntartalma, ezért folyamatos ellátása létfontosságú. Az oxigénnek a test sejtjeihez való hozzáférésének megszűnése halálához vezet. Az anyagok oxidációja során képződött szén-dioxidot el kell távolítani a testből, mivel jelentős mennyiségben történő felhalmozódása életveszélyes. Az oxigén és a szén-dioxid cseréjét a test és a környezet között légzésnek nevezzük. Emberekben és magasabb állatokban a légzési folyamat a következő sorrendben zajlik: légkörcsere a légkör és a tüdőalveolák között, gázcserék a tüdőalveolák és a vér között (külső légzés), vérgázszállítás, gázcsere a vér és a szövetek között (belső, szöveti légzés). A légzőszervek közé tartozik a légutak és a tüdő. Az orr, orrdugász, gég, légcső, hörgők és hörgők azért vezetnek levegőt a tüdő alveolusaiba, ahol gázt cserélnek.
Orrüreg
A légzőrendszer az orrüreggel kezdődik, amelyet a koponya és a porc arcrészének csontok képeznek. Az orrlyukak az orrüreg bejárataként szolgálnak, a lyukak, amelyek az üreggel kapcsolatban vannak az orrdugással, a choana kijáratává válnak. Az orrüreg falai egyenetlen megkönnyebbülést képeznek, amelynek következtében a levegőnek a nyálkahártya sejtjeinek külső rétegével történő érintkezési területe megnő. Sokan közülük van cilia. Az orrüregben áthaladó belélegzett levegőt a membránon áthatoló számos edényen átáramló vér melegíti fel, és emellett, érintkezve a nyálkahártyával, megnedvesítik és részben megtisztítják a portól, baktériumoktól és számos más szennyeződéstől. Az orrüregből a levegő bejut az orrdugárba, majd a garat orális részébe, majd a gégébe.
gége
A gége komplex felépítésű, mivel nemcsak a levegővezetésre, hanem a hangok kialakítására is szolgál. A gége különböző formájú porcokból áll, amelyeket szalagok és ízületek kötik össze, izmokat hajtva. A gége vázát pár nélkül (pajzsmirigy, krikoid és epiglottus) és párosított (arytenoid, kürt alakú és sphenoid) porcok alkotják. A legnagyobb - a pajzsmirigy porc - elöl található. A gégüreg egy nyálkahártyával van bélelt, amely a hangszálakat képezi. A szalagok szabad szélei között a glottis hosszirányában helyezkedik el.
A hangkábelek feszültségét és relaxációját speciális izmok szabályozzák. Nyugodt állapotban, amikor egy személy csendben van, a glottis nyitva van és egyenlő szárú háromszögnek tűnik. Beszélgetés vagy éneklés közben a hangkábeleket meghúzzák, összehúzzák és rezegnek, amikor a kilégzett levegő áthalad és hangot generál. A végső hangképződés azonban a szájban, az orrban, a garatban történik, és a nyelv, az ajkak, az alsó áll állásától függ. A hangmagasságot a hangkábelek hossza határozza meg: minél hosszabb a zsinórok, annál alacsonyabb a rezgés gyakorisága és annál alacsonyabb a hang. A gég bejáratát az epiglotus borítja, amely megakadályozza az étel bejutását a légzőrendszerbe. A gége leül a légcsőbe (légzési torok).
légcső
A felnőttkorban a légcső 10–13 cm hosszú cső alakú, és arra szolgál, hogy a levegő a tüdőbe és a hátba kerüljön. A hialin porc 16-20 félgyűrűjéből áll, amelyek merevséget adnak és megakadályozzák a légcső leesését. A porcos félgyűrűket egymás között sűrű kötőszövet köti össze. A félgyűrűk végei között a hát a kötőszövet membránja. A félgyűrűk közötti kötőszövetben elasztikus szálak miatt a légcső meghosszabbodhat, ha a gége felfelé mozog, és lerövidülhet, amikor leereszkedik. A légcső ürege csomózott hámmal van bélelt, amelynek ciliái a levegővel együtt eső porrészecskéket és a nyálkat a torokig vezetik, ahol azokat lenyelik. A légcső alsó vége két vékonyabb csőre oszlik - a hörgőkre (jobb és bal). Az osztódási helyet tracheális elágazásnak nevezzük.
hörgőket
A hörgők fokozatosan kisebbre ágazódnak, elérve a legvékonyabb és legvékonyabb ágakat - hörgőket, amelyek átmérője nem haladja meg a milliméter töredékét. Általában a hörgők elágazása sűrű hálózatot képez - a hörgőfát. A nagy hörgők, akárcsak a légcső, porcgyűrűkből állnak, amelyeket egy kötőszövet köt össze. A hörgőkben a porcos csontváz hiányzik, azonban nem esik le a falról, mivel simaizomszálakból áll. A bronchiók a légutak utolsó elemei.
tüdő
A tüdő kúpos alakú, páros szivacsos szervek. A tüdőszövet bronchiókból és sok apró pulmonális vezikulából - alveolákból áll -, amelyek úgy néznek ki, mint a bronchioles félgömb alakú kiemelkedései. Az alveolák falai egyrétegű hámsejtekből állnak, amelyeket egy sűrű vérkapilláris hálózat vesz körül. A Nutria-tól az alveolákat egy folyékony felületaktív anyag (boros felületaktív anyag) borítja, amely gyengíti a felületi feszültséget, és megakadályozza, hogy az alveolák teljesen kilökjenek a kilégzés során. Az alveolák és a kapillárisok teljes falvastagsága több mikrométer. Ennek a szerkezetnek köszönhetően az oxigén könnyen behatol az alveoláris levegőből a vérbe, a szén-dioxid pedig a vérből az alveolusokba. Az alveolák átmérője átlagosan 0,3 mm, de mivel az alveolák tüdeiben akár 300 millió is lehet, felnőtt felületük pedig 50–100 négyzetméter, a tüdőben a gázcsere rendkívül gyors. A tüdő (jobb és bal) a mellkasban helyezkedik el, és szorosan szomszédos a falán. A tüdő felületét egy speciális héj - pleura borítja, amely két levélből áll: a külső levél béleli a mellkas belső felületét, és a belső lefedi a tüdő felületét. A levelek között egy hermetikusan zárt résszerű hely van, amelyet pleurális üregnek hívnak. Kis mennyiségű folyadékot tartalmaz, amely hidratálja a mellhártyát, és segít megcsúsztatni őket egymáshoz képest.
A chordate főbb átalakulása a légzőrendszerben.
A légzőrendszer fejlődése.
A légzőrendszer elvégzi a test és a környezet közötti gázcserét.
Alacsony anyagcserével rendelkező állatokban a gázcsere diffúzióval történik a test teljes felületén. A testméret növekedése, a szervezetben zajló anyagcsere-folyamatok intenzitásának növekedése egy speciális légzőkészülék kifejlesztéséhez vezetett.
Az akkordok légzőrendszerének evolúciós transzformációi.
A légzés fő funkciójának erősítése:
1. a gázcsere felületének növekedése;
2. a levegő- és légzőrendszerek megkülönböztetése;
3. a légzőszervek javítása: a mellkas megjelenése
sejtek, légző izmok.
2. Az elvégzett funkciók számának bővítése: tisztítás, melegítés, párásítás; hőszabályozás, hanggenerálás.
3. Funkciók helyettesítése: a földi gerinceseknél kopoltyúkkal történő légzést a tüdőben lévő gázcserével helyettesítik.
4. Funkcióváltás: az ősi pontyok úszóhólyagját légzőszervré alakítják.
5. Funkciók és szervek szétválasztása:
1. szárazföldi gerincesekben a légzőrendszer elválasztása a
elsődleges emésztőcső;
2. a gerinces tüdőben a levegő és a légzőrendszer elválasztása.
Az evolúció során kialakul a morfofunkcionális kapcsolat a légzőrendszer és a keringési rendszerek között (dinamikus koordináció), amely kifejezésre kerül a légzőrendszerben lévő edények elhelyezkedésében, az alveolák és a kapillárisok falának felépítésében (aerogematikus gát), a gázátviteli transzportrendszerek jelenlétében és a légző pigmentek jelenlétében a vérben.
Az idegrendszer funkciói.
Az idegrendszer fő funkciója a test külső hatásainak bevezetése, amelyet az emberi test adaptív reakciója kísér. Az agy egy törzsből és az agyból áll. Az agy minden része felelős bizonyos feladatok elvégzéséért. Vegye figyelembe a központi idegrendszer funkcióit:
Mivel az előagy fel van osztva a végsőre és a közbensőre, ezért mindegyik önmagában is ellát valamilyen funkciót. Tehát a hipotalamusz, a talamusz és a limbikus rendszer a közbenső termék részét képezi. Az első a létfontosságú szükségletek (libidó, éhezés), érzelmek központja. A talamusz végzi az információ elsődleges feldolgozását, szűrését. A limbikus rendszer felelős az egyén érzelmileg impulzív viselkedéséért.
Ez az idegrendszer neuroglia nevű sejteket tartalmaz. Támogató funkciót látnak el, részt vesznek az idegrendszer sejtjeinek metabolizmusában.
A gerincvelőben fehér anyag található, amely képezi az útvonalakat. Összekapcsolják a gerincvelőt és az agyat, az agy különálló szegmenseit. Az útvonalak vezetői, reflex funkciót hajtanak végre.
Az elemzők a reflektorok szerepét játszják a külső anyagi világ emberi tudatában.
Az agykéreg aktivitása erősebb idegi aktivitást jelent, és kondicionált reflex funkciót lát el.
A központi idegrendszer fő funkciója az egyszerű és komplex reflexeknek nevezett reflektív reakciók végrehajtása.
A központi idegrendszer összeköti a központi idegrendszert a végtagokkal és a szervekkel. Csontok nem védik, ami arra utal, hogy toxinoknak és mechanikai károsodásoknak lehet kitéve.
A perifériás idegrendszer funkciói
A PNS vegetatív és szomatikus, amelyek mindegyike bizonyos funkciókat lát el. A szomatikus idegrendszer felelős a mozgások koordinálásáért és a külvilágból származó ingerek fogadásáért. Szabályozza az emberi elmét ellenőrző tevékenységeket.
A vegetatív viszont védő funkciót lát el veszély vagy stresszes helyzet esetén. Felelős a vérnyomásért és a pulzusért. Amikor valaki aggódik, az izgalom érzékeltetése után növeli az adrenalin szintjét.
Az autonóm részét képező parasimpátikus rendszer akkor látja el funkcióit, amikor az egyén nyugalomban van. Felelős a pupillák szűkítéséért, az urogenitális és az emésztőrendszer stimulálásáért.
És mégis, milyen funkciókat lát el az idegrendszer?
Információszerzés az emberi világról és a test állapotáról.
Ezen információk átvitele az agyra.
A test központi idegrendszerének tudatos működésének koordinálása.
A pulzusszám, a hőmérséklet stb. Koordinálása és szabályozása
Az agy különféle részeinek szerkezete és funkciói a halakban, kétéltűekben, hüllőkben, madarakban, emlősökben és emberekben.
Az ichthyopsid, a sauropsid és az emlős típusú agyok közötti különbségek.
Az idegrendszer fejlődésének fő irányai a gerincesek különböző képviselőiben.
Az agy fő részeinek homológiája a gerincesek különböző képviselőiben.
Terminális teratogén tényező.
Onfilogenetikusan okozta rendellenességek a légzőrendszerben az emberekben. Példák.
Az agy és a gerincvelő fő veleszületett filogenetikailag meghatározott rendellenességei az emberekben. Példák.
A földi hozzáférés és a magasabb gerincesek számára jellemző aktívabb élet miatt a hüllő agyának minden része fokozatosan fejlődik.
1. Az előagy jelentősen meghaladja a többi osztályt. A köpeny vékony marad, de felületén az idegsejtek mediális és oldalsó felhalmozódása jelenik meg - ez egy olyan szürke anyag, amely az agyfélteke embrionális kéregét képviseli. A hüllőben a kéreg még nem játszik szerepet az agy felső részében, hanem a legmagasabb szaglási központ. A filogenezis folyamatában azonban a szagláson túl egyre növekvő és más típusú érzékenységet is felvetett az emlősök agykéregének megjelenéséhez. A hüllők előagyjának félteke teljesen lefedi a diencephalont. A magasabb szintű integráló központ szerepét a csíkos test végzi (agy sauropsy típusa)
2. A diencephalont a látógumók és az algumók régió képezik. A hátsó oldalán a tobozmirigy és egy speciális parietális szerv található, amelynek a gyíkok szemszerű felépítése van. A ventrális oldalon az agyalapi mirigy található.
3. A középső agy meglehetősen nagy, kétrészes megjelenésű. Ez a vizuális észlelés központja, amely nagy jelentőséggel bír a szárazföldi állatok számára.
4. A kisagy félkör alakú lemeznek tűnik, rosszul fejlett, de jobb, mint a kétéltűek, a mozgások összehangolásának komplikációja miatt.
5. A medulla oblongata éles hajlítást képez, mely a magasabb gerincesekre jellemző. Magjaiból származnak a koponya idegei.
Összesen a hüllőknek 12 pár agyidege van.
Agy

Gerinces agy evolúciója: a - hal; b - kétéltű; c - hüllő; g jelentése emlős; 1 - szaglási lebenyek; 2 - az agy; 3 - középső agy; 4 - kisagy; 5 - a medulla oblongata; 6 - diencephalon
1. Az előagy jól fejlett, a félteke jelentős, részben fedezi a diencephalont. A félgömbök növekedése azonban a nyálkahártya (az agy sauropszia típusa) fejlődése miatt következik be, nem pedig a kéreg. A szagló lebenyek nagyon kicsik, mivel a szaglás érzéke elveszíti vezető jelentőségét.
2 A diencephalon kicsi, az agy féltekéje lefedi. A dorsalis oldalon a tobozmirigy (rosszul fejlett), a ventrális oldalon pedig az agyalapi mirigy található.
3. A középső agy meglehetősen nagy, a nagy látógörbék (dicollis) miatt, ami a látás fokozatos fejlődéséhez kapcsolódik.
4. A kisagy fejlett a mozgások repülés közbeni összetett összehangolása miatt. Keresztirányú és kéregű.
5. A medulla oblongata magok formájában lévő idegsejtek gyűjteményét tartalmazza, ahonnan az 5. és a 12. pár közötti agyidegek származnak.
Csak 12 pár agyideg.
Emlős test
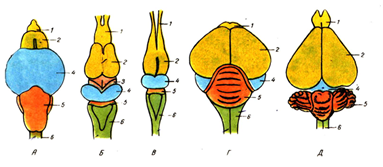
Gerinces agy evolúciója: a - hal; b - kétéltű; c - hüllő; g jelentése emlős; 1 - szaglási lebenyek; 2 - az agy; 3 - középső agy; 4 - kisagy; 5 - a medulla oblongata; 6 - diencephalon
1 Az előagy különösen nagy méretű, az agy fennmaradó részeit lefedve. Növekedése a kéreg miatt következik be, amely a nagyobb idegrendszeri tevékenység fő központjává válik (emlős agytípus). A kéreg területe növekszik a kanyarodások és barázdák kialakulása miatt. A legtöbb emlősnél az agyfélteke előtt (a cetfélék, főemlősök és - beleértve az embereket is) - nagy szaglólebeny található, ami összefüggésben van a szaglás nagy jelentőségével az állatok életében.
2 Az optikai tuberkulumok (thalamus) és az al-tubercle régió (hypothalamus) által alkotott diencephalont az előagy féltekei rejtik el. A háti oldalán a tobozmirigy, a ventrális oldalon pedig az agyalapi mirigy található.
3 A középső agyt az elülső agy félteke borítja, megkülönbözteti viszonylag kicsi méretét, és nem a kettősség, hanem a kvadrupol képviseli. A középső agy ürege, vagy a sylvian vízvezeték csak egy keskeny hézag.
4 A kisagy fejlett és bonyolultabb felépítésű; a középső részből áll - egy féreg keresztirányú barázdákkal és párosított félgömbökkel. A kisagy kialakulása a mozgások összehangolásának komplex formáit biztosítja.
5 A medulla oblongata részben a kisagy. A többi osztály képviselőitől abban különbözik, hogy az idegrostok hosszanti kötegeit - a kisagy hátulsó lábait - a negyedik kamra áramlása választja el, a hosszirányú gerincek - a piramisok - pedig az alsó felületen vannak. 12 pár agyideg távozik az agytól
48. 50. Az immunválasz filogenetikai úton megállapított típusai és formái. A gerinces immunrendszer jellemzőinek jellemzése.
Az immunrendszer filogenezise.
Az immunrendszer megvédi a testet a genetikailag idegen test behatolásától: mikroorganizmusok, vírusok, idegen sejtek, idegen testek. Tevékenysége azon a képességén alapszik, hogy meg tudja különböztetni saját struktúráját a genetikailag idegentől, megsemmisítve azt.
Az evolúció során az immunválasz három fő formája alakult ki:
1) 1. Phagocytosis vagy idegen anyag nem specifikus megsemmisítése;
2) 2. Sejtes immunitás, amelynek alapja az ilyen anyag T-limfociták általi specifikus felismerése és megsemmisítése;
3) 3. Humorális immunitás, amelyet B-limfociták leszármazottjainak, az immunglobulinok úgynevezett plazmasejtjeinek és az idegen antigének kötődésének köszönhetően hajtanak végre.
Az evolúció során az immunválasz kialakulásának három szakaszát lehet megkülönböztetni:
1. 1. Kvázi immunitás (lat like) felismerés saját és idegen sejtek organizmusai. Az ilyen típusú reakciókat bél és emlősök között figyeljük meg. Ez a reakció nem kapcsolódik az immuntestek előállításához, és ugyanakkor nem alakul ki immunmemória, vagyis az idegen anyag újbóli behatolásakor még nem erősödik az immunválasz.
2. 2. Primitív sejtes immunitás megtalálható annelidekben és tüskésbőrben. Ezt coelomocyták - a test másodlagos üregének sejtjei - biztosítják, amelyek idegen anyagokat tudnak elpusztítani. Ebben a szakaszban immunológiai memória jelenik meg.
3. 3. Integrált sejtes és humorális immunrendszer. Jellemzője az idegen testekkel szembeni specifikus celluláris és humorális reakciók, az immunrendszer limfoid szerveinek jelenléte és antitestek képződése. Ez a fajta immunrendszer nem jellemző a gerinctelenekre.
A cyclostomata antitesteket képezhet, de továbbra is nyitott a kérdés, vajon thymus mirigyük van-e, mint az immunogenezis központi szerve. A thymusot először a halakban észlelik.
Az emlősök nyirokhártya szerveinek evolúciós prekurzorai - thymus, lép, a lymphoid szövetek felhalmozódása - teljes egészében kétéltűekben található meg. Az alsó gerincesekben (halak, kétéltűek) a csecsemőmirigy aktívan enged fel antitesteket, ami jellemző a madarakra és az emlősökre.
A madarak immunválaszának sajátossága a limfoid szerv mintájának - egy gyári zsáknak - a jelenléte. Ebben a szervben B-limfociták képződnek, amelyek antigén stimuláció után képesek plazma sejtekké alakulni és ellenanyagokat termelni.
Az emlősökben az immunrendszer szervei két típusra oszthatók: központi és perifériás. A központi szervekben a limfociták érése az antigének jelentős hatása nélkül történik. A perifériás szervek fejlődése éppen ellenkezőleg, közvetlenül az antigénhatástól függ - csak az antigénnel való érintkezéskor kezdődnek meg a limfociták szaporodási és differenciálódási folyamatai.
Az emlősök immunogenezisének központi szervei a thymus, ahol T-limfociták képződnek és szaporodnak, valamint a vörös csontvelő, ahol B-limfociták képződnek és szaporodnak.
Az embriógenezis és a tojássárgáj korai szakaszában az őssejtek nyálkahártyái és a vörös csontvelőbe vándorolnak. Születés után a vörös csontvelő az őssejtek forrásává válik.
A perifériás limfoid szervek a következők: nyirokcsomók, lép, mandula, a bél limfoid tüszői. Születésükre még nem alakultak ki, és a limfociták képződése csak antigén stimuláció után kezdődik meg, miután az immunogenezis központi szerveiből származó T és B limfociták betelepítették őket.
49. 51. Ontogenezis, típusai és periodizációja.
egyedfejlődés, vagy az egyéni fejlődés: a testben a zigóta kialakulásának pillanatától a halálig zajló átalakulások halmaza. A „ontogenezis” kifejezést E. Haeckel biológus kezdte először 1866-ban (a görög onszubstanciáról és a genezis fejlődéséről).
A ontogenezis doktrínája - Ez a biológia egyik ága, amely a szervezetek egyéni fejlődésének mechanizmusait, szabályozását és jellemzőit vizsgálja.
A ontogenezis ismerete nemcsak általános elméleti jelentőséggel bír. Az orvosoknak meg kell ismerniük a különböző életkori kóros folyamatok sajátosságait, meg kell akadályozniuk a betegségeket, és meg kell oldaniuk a különféle korcsoportok számára a munka és a szabadidő megszervezésével kapcsolatos társadalmi és higiéniai problémákat.
Kétféle ongenezis létezik: közvetett és közvetlen. közvetett lárva formájában halad tovább. A lárvák aktív életmódot folytatnak, ők maguk is keresnek ételt. A lárvák életfunkcióinak megvalósításához számos átmeneti (átmeneti) szerv hiányzik a felnőtt szervezetekben. Az ilyen típusú fejlődést metamorfózis (átalakulás), a test anatómiai és élettani átalakulása kíséri. Ez jellemző a gerinctelenek (szivacsok, bél, férgek, rovarok) és leszálló gerincesek (kétéltűek) különböző csoportjaira.
Közvetlen fejlesztés előfordulhat nem lárva formában, vagy intrauterin lehet. Nem lárva típusú A fejlődés halakban, hüllőkben, madarakban, valamint gerinctelenekben fordul elő, amelyek tojásban gazdag tojássárgája - ez egy tápanyag, amely elegendő a ontogenezis teljesítéséhez. Táplálás, légzés és ürítés céljából az embriókban az embrionális szervek is kialakulnak.
Intrauterin típusú emlősökre és emberekre jellemző fejlődés. Tojásuk szinte semmilyen tápanyagot nem tartalmaz, és az összes létfontosságú funkciót az anya testén keresztül végzik. E tekintetben az embriók ideiglenes szervekkel rendelkeznek - az embrionális membránok és a méhlepény, amely kapcsolatot teremt az anya és a magzat között. Ez a filogenezis legújabb típusa az ontogenezisben, és biztosítja az embriók legjobb túlélését.
Az ontogenezis számos egymást követő és a genetikailag programozott periódus fő jellemzőit foglalja magában:
1. Preembrionális (más néven proembrionális vagy pre-zigotikus időszak, vagy progenesis);
2. Az embrionális (vagy embereken szülés előtt) időszak;
3. Postembryonic (vagy postnatalis ember számára) időszak.
a. 52. Az pre-zigotikus időszak általános jellemzői, az embrionális fejlődés stádiuma. Kritikus időszakok. Teratogén tényezők.
KÉSZÍTETT IDŐSZAK
Ez a periódus a szülők testében fordul elő, és kifejeződik a gametogenezisben - az érett tojások és sperma kialakulásában.
Jelenleg ismert, hogy ebben az időszakban számos olyan folyamat zajlik, amely közvetlenül kapcsolódik az embrionális fejlődés korai szakaszaihoz. Tehát a tojások érlelése során a pakhinemben megfigyelt meiosis volt gén amplifikáció (számos másolat létrehozása), amelyek felelősek az r-RNS szintéziséért, amelyet izolálnak a DNS-től és felhalmozódnak a nukleoliák körül. Ezeket a géneket bevonják a transzkripcióba az embriogenezis korai szakaszában, biztosítva a riboszómák kialakulásában résztvevő r-RNS felhalmozódását. Ezen túlmenően a pre-zigotikus időszakban az i-RNS felhalmozódik a jövőbeni felhasználáshoz, amely csak a zigóta fragmentáció korai szakaszában kerül be a protein bioszintézisbe.
A tojásokban az ovogenezis során felhalmozódik sárgája, glikogén és zsírok, amelyeket az embriogenezis folyamatában fogyasztanak.
A tojássárgája tartalom alapján (lecitos) petesejt lehet:
Oligolecitic (alacsony sárgája);
Mesolecitic (átlagos sárgája mennyiségével);
· Poliecit (több sárgája).
A tojássárgája eloszlásának jellege szerint a tojás citoplazmájában:
· Izolecital (görög izosz - egyenlő, a sárgája egyenletesen oszlik el a sejtben);
· Telolecital (görög thelos - a vége, a sárgáját közelebb tolják a vegetatív pólushoz, a sejtmag pedig közelebb van az állathoz);
Centrocytal (a tojássárgája a tojás középső részén helyezkedik el)
Az izolecitalis sejtek jellemzőek a lándzsára és az emlősökre, telolecital - kétéltűekre (közepesen telolecital, hüllőkre és madarakra - élesen telolecitalis), centriocitális - rovarokra.
Egyes állatfajok petesejtjei már a megtermékenyítés előtt is kétoldalú szimmetriát kapnak, azonban ez továbbra is instabil, és később átfordulhat.
Sok állatfajban, még a megtermékenyülés megkezdése előtt elkülönítésaz organellák (újraelosztás) és a petesejtbe záródások; felhalmozódik a glikogén és az RNS az állati póluson, a Golgi-komplex és az aszkorbinsav az Egyenlítőn. A szegregáció a megtermékenyítés után folytatódik.
EMBRYONIAL IDŐSZAK
Az embrionális periódus egy zigótával kezdődik, vagy akár fiatal egyedeknek a tojásmembránokból történő felszabadulásával, vagy egy új szervezet megszületésével ér véget. Ez az időszak a következő szakaszokból áll: zigóták, zúzás, gasztronáció és hiszto- és organogenezis.
AZ EMBRONICON FEJLESZTÉSI FÁNYOK JELLEMZŐI
Ember példájában.
A megtermékenyítés után megtörténik az embrionális fejlődés első stádiuma - a zigóta stádium (egysejtű embrió stádiuma). A zigóta, mivel egy sejt, képes holisztikus többsejtű organizmus kialakulására, azaz teljes potenciállal rendelkezik.
Szakasz zúzás: ettől a stádiumtól kezdve az embrió többsejtűvé válik, de gyakorlatilag nem haladja meg a zigóta méretét. A zúzás az, hogy bár a sejtek mitózissal osztódnak, nem növekednek az anyai sejtek méretéig, mert hiányzik heteroszintetikus interfázis, és az autoszintézis interfázis G1 periódusa az előző osztódás telofázisában fordul elő. A zúzási szakasz egy blastula képződésével ér véget. Az első blastomer, mint a zigóta, a totipotencia tulajdonságával rendelkezik, amely alapja a monozigótikus (azonos) ikrek születésének.
Az emberekben a blastula a fejlődés 6-7. Napján képződik, és buborékának (blastocisztta) jelenik meg, amelynek falait egyetlen sejtréteg alkotja - trofoblaszt, amelyek ellátják a táplálkozást és a kiválasztást. A vezikuláció belsejében felhalmozódnak a sejtek - egy embrióblast, amelyből az embriótest tovább fejlődik.
A színpadon gasztruláció (7–19 napos személynél) csírarétegek (ektoderma, endoderma és mezoderma) képződése megtörténik, és axiális szervek (akkord, idegi cső és bélcső) komplexét fektetik.
Az időszakban hisztogenezis és organogenezis van egy könyvjelző az ideiglenes (ideiglenes) és a végleges (végleges) szervekről. A gerincesekben, beleértve az embereket, ideiglenes szerveket hívnak csíramembránok. A tojássárgáj kifejlődése minden gerincesre jellemző. Halakban, kétéltűekben, hüllőkben és madarakban sárgáját tartalmaz, és trofikus és hematopoietikus funkciókat lát el. Valódi szárazföldi állatokban a tojássárgája mellett egy folyadékkal töltött amnion is található, amely vizes közeget hoz létre az embrió fejlődéséhez. Amnion gerinceseket (hüllők, madarak és emlősök) hívnak magzatburkosokde nincs meg - anamniotes (halak, kétéltűek).
Hüllőknél és madaraknál a tojássárgája és az amnion mellett a következőket fektetik le: allantois (húgyzsák, amely karbamidot halmoz fel) és a seros membrán (biztosítja az embrió légzését). Az emlősökben a serozus membrán helyett chorion (zavaros membrán) képződik, amely táplálékot, légzést és ürítést biztosít a magzat számára. A koriont trofoblasztból és kötőszövetből képezik. A placentació stádiumától kezdve részt vesz a placenta kialakulásában. Az amnion amniotikus folyadékot tartalmaz. Az elsõ erek és az elsõ vérsejtek a sárgája zsákban alakulnak ki. Az emlősökben és emberekben az allantois meghatározza a placenta helyét.
A hiszto- és organogenezis az emberekben kezdődik negyedik hét és születéskor ér véget.
Az első az úgynevezett elsődleges ektoderma a sejteket elválasztják, idegi lemezt képezve, amelyből az idegrendszer összes szerve és az érzékszervek egy része később fejlődik ki. A fennmaradóból másodlagos ektoderma az epidermiszt és annak származékait lerakják - faggyú, izzadság, emlőmirigyek, körmök, haj és néhány egyéb képződmény.
-tól endoderma kialakult: a gyomor-bél traktus, légzőrendszer, máj és hasnyálmirigy hámja.
-tól mezodermából - csontváz, csíkos és simaizmok, szív- és érrendszer, valamint az urogenitális rendszer fő része.
KRITIKAI FEJLESZTÉSI IDŐSZAKOK
1921-ben a Stockcard C.R. megalapozta az állati organizmusok úgynevezett kritikus fejlődési periódusaival kapcsolatos ötleteket. Később hazánkban Svetlov P. G. foglalkozott ezzel a problémával, aki 1960-ban megfogalmazta a kritikus fejlődési periódusok elméletét és kísérletileg igazolta. Lényege abban rejlik, hogy az embrió fejlődésének minden stádiuma egy kvalitatív új szerkezetátalakítás rövid időszakával kezdődik, amelyet a sejtek meghatározása, proliferációja és differenciálódása kísér. Ebben az időszakban a különféle káros környezeti tényezők - fizikai, kémiai és egyes esetekben - biológiai is különösen érzékenyek, amelyek felgyorsíthatják, lelassíthatják, sőt megállíthatják a fejlődést.
A humán ontogenezisben a következő kritikus periódusokat különböztetjük meg: 1) gametogenezis; 2) megtermékenyítés; 3) implantáció; 4) axiális szervek komplexének kialakulása és a placenta kialakulása (3-8 hét) 5) egy adott szerv vagy szervrendszer differenciálódási periódusai (20-24 hét); 6) születés; 7) az újszülött periódusa (legfeljebb 1 év); 8) pubertás.


b. 53. Az embriógenezis fő mechanizmusai.
AZ EMBRIOGENÉZIS ÁLTALÁNOS MECHANIZMUSAI
1. Sejtosztódás
2. A sejtek differenciálódása
3. Differenciális génaktivitás
4. Magzati indukció
5. Intercelluláris interakciók
6 A sejtek migrációja.
7. Sejthalál
8. A fejlődés klón alapelve
9. Növekedés.
10. Morfogenezis.
Az embrionális fejlődés különféle folyamatokon / mechanizmusokon alapszik, amelyek magukban foglalják: sejtosztódást, differenciálódást, embrionális indukciót, intercelluláris kölcsönhatásokat, sejtek vándorlását, sejthalálát, a klonális fejlődés elvét, növekedést, morfogenezist és differenciált génaktivitást.
1. Sejtosztódás megalapozó sejtburjánzás / sejtnövekedés / és ez a fő mechanizmus a növekedés biztosításához, vagyis a testtömeg és a méret növekedéséhez. Ezenkívül a sejtosztódás során számos esetben a genetikai programokat átváltják, és ennek eredményeként a sejtek bizonyos funkciók elvégzésére szakosodtak.
2. A sejtek differenciálódása - Ez egy folyamat, amikor a speciális sejtek látszólag egységes sejtekből és komplexeikből származnak, amelyek különböznek az anyai morfológiai és funkcionális tulajdonságaitól. Ez a folyamat divergens / többirányú / karakterű. Biokémiai szempontból a differenciálás bármelyik lehetséges bioszintézis útvonal közül választhat (például az eritrocita prekurzor sejtek választják a hemoglobin szintézis útvonalat, a kristályos lencsesejtek pedig a kristályos fehérjét). Morfológiai szempontból a differenciálódást kifejezik a specifikus szerkezeti jellemzők megszerzésében.
A differenciálódás eredményeként olyan magasan specializálódott sejtpopuláció alakul ki, amely elvesztette a magját (eritrociták, keratinizált epidermális sejtek), vagy a sejtekben megindul a nagyon specifikus anyagok szintézise, \u200b\u200bpéldául az aktin és a miozin összehúzódó fehérjéi - az izomrostokban, bizonyos hormonok - az endokrin mirigyek sejtjeiben stb. d.
Az egyes sejtek differenciálódásának útját genetikailag meghatározzuk / előre meghatározzuk /. A zúzás szakaszában a sejtek meghatározása továbbra sem stabil (labilis), és a differenciálódás iránya megváltozhat. Ezt a huszadik század első negyedévében Hans Spemann megerősítette az ászok kísérletein. Az egyik faj tritonjából vett ektodermális sejteket átültette a másik endodermájába. Noha a donor sejtek színben különböztek a recipiens sejtektől, ugyanolyan alulmentekben fejlődtek ki, mint a körülvevő recipiens sejtek. Ha a donorok olyan szervezetek voltak, amelyek befejezték a gastrulációs folyamatot, akkor a bőrbe átültetett neurális lemez ektodermális sejtjei az idegszövet rügyéhez vezettek, azaz differenciálódásuk útja már előre volt meghatározva.
Most ismertek olyan tényezők, amelyek meghatározzák a szövetek differenciálódását. A legkorábbi tényező, amely már a blastula szakaszában megjelenik elkülönítés/ lat A zigóta „szétválasztása” / citoplazmatikus struktúrái, amelyek miatt zúzás közben a citoplazma különböző szakaszai esnek az első robbanásgomba. Tehát nyilvánvalóan az oociták citoplazmájának különböző részeiben létező jelentéktelen minőségi különbségek befolyásolják a blastomerek sorsát. Bizonyítékok vannak arra is, hogy sok embrionális szövet differenciálódása csak bizonyos kritikus számú sejt jelenlétében fordulhat elő.
A sejtek differenciálódásának fő mechanizmusa a gének differenciál aktivitása.
3. Magzati indukció - ez az egyik szövet vagy egy embriószerv / induktor embriójának / más szervembriók fektetésének a hatása. Tehát például gerinces állatokon a chordo-mesodermal komplex elrendezése indukálja / indukálja az idegcső fejlődését / fektetését.
A indukáló hatások másik formája az intercelluláris kölcsönhatások.
4. Intercelluláris interakciók hézagcsomópontokon keresztül hajtják végre, ahol egyes sejtek plazmamembránja szorosan érintkezésbe kerül más sejtek plazmalemmájával. Ezen kapcsolatok területén gyenge elektromos áram, szervetlen anyagok ionjai vagy akár viszonylag nagy szerves anyagok molekulái továbbadhatók a sejtek között.
5 A sejtek migrációja. Az embriógenezis során mind az egyes sejtek, mind azok komplexei különböző távolságra migrálnak. Az egyes sejtek általában amőboid mozgással vándorolnak, miközben folyamatosan vizsgálják a környezetüket.
6. Sejthalál (apoptózis) az embrió fejlődésének számos szakaszában szükséges folyamat. Tehát a lábujjak és a kezek elválasztását az interdigitális terekben található sejtek elpusztítása előzi meg.
7. A fejlődés klón alapelve. Kísérletileg kimutatták, hogy a korai embrió sok sejtjének nem célja a további fejlődésben való részvétel. Számos embrionális struktúra olyan sejtekből épül fel, amelyek csak egyetlen, kis számú sejt megosztásakor alakulnak ki.
8. Növekedés. A növekedés alatt a testtömeg és méretének növekedését értjük. A növekedés egyenetlen, az egyes szövetek és az embrió különböző részei eltérő sebességgel növekednek.
9. Morfogenezis. Ez az embrió testrészeinek és szerveinek külső és belső konfigurációjának térbeli kialakulásának folyamata. Nincs egy általánosan elfogadott elmélet, amely magyarázza ennek a folyamatnak a mechanizmusát. A legalkalmasabb helyzeti információ fogalmaL. Volpert által javasolt / 1975 / szerint, amely szerint a sejtek képesek érzékelni a helyzetinformációkat, amelyek tartalmazzák a sejtek más sejtekhez viszonyított helyének jelzését, és ezáltal meghatározzák azt a tervet, amely szerint az embrió fejlődése zajlik.