Обоняние. В полости носа имеются рецепторы органа обоняния.
Выделение. Некоторые вещества (продукты жизнедеятельности и т.п.) могут выделяться через дыхательную систему.
Защитная. Имеется значительное количество специфических и неспецифических иммунных образований.
Регуляция гемодинамики. Легкие при вдохе усиливают приток венозной крови к сердцу.
К сожалению, доказательств, которые касаются этого последнего вопроса, непросто. Тем не менее, единственный вывод о каком-то значении касается вокализации у птиц. Вокализация у птиц, как и у человека, контролируется аппаратом, который не требует самостоятельных движений в левой и правой половине.
Может заключаться в том, что отсутствие необходимости в равном, но независимом представлении двух половин речевого аппарата является важным фактором в эволюции асимметрий головного мозга человека, поскольку аналогичные асимметрии возникают у песенных птиц. У изученных птиц орган звукоизвлечения, сиринкс, имеет мышцы с левой и правой сторон, и каждый набор реагирует на отдельную ветвь гипоглобулярного черепного нерва. Левый набор сиринковых мышц значительно больше правого, что само по себе наводит на размышления, но четкие результаты достигаются путем отделения нерва от левых мышц и сравнения эффектов этой операции с эффектами резания нерва к мышцам на правой стороне сиринкса.
Депо крови.
Терморегуляция.
Особенности строения органов дыхания у рыб, амфибий, рептилий, птиц, млекопитающих и человека.
Особенности дыхательной системы круглоротых, хрящевых, костных рыб
В отличие от всех других позвоночных у круглоротых в жаберных щелях развиваются жаберные мешки, имеющие энтодермальное происхождение. Внутренняя поверхность жаберных мешков образует многочисленные складки слизистой оболочки, в которых расположена густая сеть мелких кровеносных сосудов - капилляров. У миноги каждый жаберный мешок (всего их 7 пар) открывается наружу самостоятельным наружным отверстием.
Если разрез слева, все или почти все слоги, ранее использовавшиеся канарейкой, теряются, тогда как если мышцы правой стороны инактивируются, вокализация даже сразу после этого практически не изменяется, причем только один или два слога отсутствуют большинство.
Так как в канарейке левый гипоглоссальный нерв исходит из левой части мозга, можно предположить, что лево-мозговая доминантность вокализации у канарейки, а другие эксперименты подтверждают, что это так. У человека есть область лобной доли левого полушария, которая называется площадью Броки, потому что французский невролог этого имени обнаружил, что пациенты, у которых он был поврежден, страдали формой афазии. Он расположен во внешнем полосатом теле, который по другим причинам считается аналогичным коре головного мозга млекопитающих.
Внутреннее отверстие жаберного мешка соединяет его полость с дыхательной трубкой, которая представляет собой слепой вырост, спереди соединенный с ротовой полостью и отграниченный от нее подвижной складкой - парусом. У плавающей миноги вода поступает в ротовую полость, оттуда проходит в дыхательную трубку и, пройдя сквозь жаберные мешки, через их наружные отверстия выбрасывается наружу. При этом кислород, растворенный в воде, проникает в капилляры и связывается пигментом крови, а насыщающий венозную кровь углекислый газ переходит в воду и с ней выводится наружу.
Хотя поражения в этой области в левом полушарии привели к немедленной потере почти всей песни, поражения, помещенные в соответствующую точку в правом полушарии, позволили воспроизвести более половины обычных слогов сразу после операции в виде, который был человеческое ухо неотличимо от пред-операции. Прогноз нарушения языка, который является следствием повреждения головного мозга у пациентов с людьми, не является хорошим, но значительное выздоровление вовсе не редко или спонтанно или с помощью речевой терапии, особенно после относительно чистых повреждений из-за несчастных случаев или военных ран, а не ударов или опухоли.
В том случае, когда минога питается (или если она присосалась к какому-либо предмету), ток воды через ротовую полость становится невозможным. Парус при этом закрывает вход в дыхательную трубку, не пропуская воду, а жидкая пища проходит из ротовой полости в пищевод. При этом дыхание осуществляется иным путем: под воздействием мускулатуры стенок тела жаберные мешки сжимаются, и вода выталкивается через наружные жаберные отверстия (активный выдох); далее, благодаря упругости хрящевой жаберной решетки жаберная область снова расширяется, и вода через те же наружные отверстия вновь засасывается в жаберные мешки (пассивный вдох).
Другая параллель с механизмами человеческого языка заключается в том, что если левая ветвь гипоглобулярного нерва, идущая к сиринкс разрезан молодыми птицами, правые структуры очень быстро берут на себя контроль над песней. Является ли слуховое восприятие или понимание звуков столь же латеральным в мозге песни-птицы, как кажется, остается вокальное производство Акустическая латерализация совершенно другого вида известна у нескольких видов сов, некоторые из которых имеют очень выраженные физические асимметрии внешнего уха, последовательно у всех индивидуумов.
Основную часть дыхательной системы хрящевых рыб составляют большие жаберные пластины эктодермального происхождения, прикрепленные одной из сторон к межжаберным перегородкам. При вдохе глотка расширяется, вода поступает через рот и омывает жаберные пластины. При выдохе объем глотки уменьшается и вода выходит наружу через жаберные щели, их бывает 5-7. Жаберных крышек у хрящевых рыб нет. Кроме жаберных щелей у большинства видов сзади глаз есть рудиментарные жаберные щели - брызгальца, открывающиеся в переднюю часть глотки, куда через них поступает вода при вдохе.
Было бы любопытно, действительно ли из всех позвоночных только песенных птиц, сов и людей воспользовались релаксацией двусторонней симметрии мозга. К сожалению, до сих пор неясно, является ли латеральная функция человеческого мозга предвестником других приматов, и это, очевидно, является решающим моментом. В терминах анатомических неравенств Каннингем обнаружил, что у человеческих сильвианских трещин асимметрии были сходства у великих обезьян и крупных обезьян, и это подтвердили Йени-Комшан, Бенсон, Лемей и Гешвинд, Каин и Вада.
На мой взгляд, лучшая оценка в настоящее время заключается в том, что асимметрия функции мозга как таковой не является прерогативой человека, хотя одна из вещей, которая является латеральным языком. Латеральность церебральной области не является ни необходимым, ни достаточным условием.
Главными органами дыхательной системы костных рыб являются жабры, состоящие из многих лепестков, прикрепленных проксимальными концами к жаберным дугам в отличие от жаберных пластин хрящевых рыб, прикрепленных одной стороной к межжаберным перегородкам. Следовательно, поверхность жабр костных рыб значительно больше, чем у хрящевых рыб. Подвижная жаберная крышка прикрывает и защищает жабры.
Для человеческой умственной деятельности, и поэтому мы не должны предполагать, что степень латеральности мозга определяет степень, в которой животный вид, как говорят, обладает познаниями. Иерархический дизайн у позвоночных. Спекулятивное упрощение, разделяемое филогенетической теорией, человеческой неврологией и другими попытками дать общее описание функции мозга - это идентификация иерархии уровней активности мозга. На самом низком уровне спинной мозг явно обеспечивает самый прямой маршрут между сенсорным входом и выходом двигателя.
При вдохе ротовая полость расширяется и давление в ней падает, вследствие чего через ротовое отверстие внутрь заходит вода. Одновременно под напором воды закрывается задний конец жаберной крышки, препятствуя поступлению воды с этой стороны. В то же время, сокращаются мышцы жаберной крышки и жаберная полость расширяется. При выдохе ротовое отверстие и входное отверстие пищевода закрываются, а дно ротовой полости поднимается. В результате вода выталкивается через жаберные щели в жаберную полость и, пройдя над жабрами, выходит затем наружу около заднего края жаберной крышки, открытой теперь под напором воды.
На самом высоком уровне некоторые области коры млекопитающих получают сенсорную информацию, которая последовательно трансформируется, фильтруется и классифицируется на предыдущих стадиях, а другие области коры инициируют действия, которые могут быть даны более детальной организацией нижними отделами головного мозга.
Между спинным мозгом и корой головного мозга, среднего мозга, мозжечка, таламуса и стриатума тела могут быть назначены конкретные посреднические роли. Очевидно, что основное предположение состоит в том, что более высокие уровни организации мозга связаны со сложными формами познания и мысли, тогда как более низкие уровни касаются только более механических и рефлексивных вопросов. Во-первых, многие важные детали обязательно игнорируются: определение уровней функции делает вещи более понятными, но уровни могут в некоторых смыслах быть удобными фикциями.
Усложнение дыхательной системы
Почти у всех земноводных есть легкие. Так как внутренняя стенка легких почти гладкая, то их поверхность относительно невелика. Трахея почти неразвита, и легкие связаны непосредственно с гортанью. Т. к. грудной клетки у земноводных нет, дыхание обеспечивается работой мышц ротовой полости. При открытых ноздрях (которые в отличие от ноздрей рыб сквозные, т. е. кроме наружных ноздрей имеются и внутренние ноздри - хоаны) и закрытом рте оттягивается дно обширной ротовой полости и в нее поступает воздух. Затем ноздри закрываются особыми клапанами, дно ротовой полости поднимается, и воздух нагнетается в легкие. Выдох происходит в результате сокращения брюшной мускулатуры. Значительное количество кислорода земноводные получают через кожу и слизистые оболочки ротовой полости. У некоторых видов саламандр легких совсем нет, и весь газообмен совершается через кожу. Однако кожа только в том случае может выполнять функции дыхания, если она влажная.
Во-вторых, если производятся деления функции, почти неизбежно, что кто-то уступает искушению налагать ложные значения на дивизии. Как предостерегающий пример, даже разделение между сенсорными и моторными функциями иногда рассматривается с подозрением. Рисунок 4 Именование частей и общий план мозга позвоночных. Верхняя диаграмма представляет собой приблизительный эскиз анатомических отношений между структурами, которые обычно идентифицируются в любом мозге позвоночных. Нижняя диаграмма схематически показывает точки входа основных сенсорных нервов и сильно упрощенные пути нейронных связей в мозге.
Поэтому обитание земноводных в условиях недостаточной для них влажности воздуха невозможно. Личинки, живущие в воде, дышат жабрами (сначала - наружными, потом - внутренними) и кожей. У некоторых хвостатых земноводных, постоянно живущих в воде, жабры сохраняются всю жизнь. Таким образом, по способам дыхания амфибии еще близки к рыбам. Дыхательные пути пресмыкающихся начинаются наружными носовыми отверстиями - ноздрями. Далее воздух через носовой проход и внутренние ноздри - хоаны, попадает в ротовую полость. В глубине ротовой полости расположена гортань. Она снабжена особой мускулатурой и связана с подъязычным аппаратом. Из ротовой полости вдыхаемый воздух через гортань попадает в трахею.
У млекопитающих и птиц передний мозг больше, чем указано здесь, а мозг в целом сжимается по длинной оси. У млекопитающих постталамические проекции состоят из неокортекса полушарий головного мозга. Сложность сенсорных и моторных систем. Путем интерпретации мозговой организации с точки зрения сенсорного ввода, движущегося вверх через уровни заднего мозга, среднего мозга и переднего мозга, постепенно более отдаленные от самих органов чувств, с запланированными действиями, скоординированными последовательностями ответа и механическими рефлексами, каскадирующими назад к периферическим мышцам, один находится под угрозой чрезмерно подчеркивая простые категории ввода и вывода.
Трахея проходит вдоль шеи и в грудной полости, примерно на уровне сердца, разделяется на два коротких бронха, входящих в легкие. Легкие представляют собой тонкостенные полые мешки. По сравнению с легкими земноводных у пресмыкающихся они имеют более сложную внутреннюю структуру: внутренние их стенки, в которых ветвятся капилляры, имеют губчатое строение, что увеличивает общую дыхательную поверхность легких. Легкие - единственный орган дыхания пресмыкающихся. Кожа этих животных сухая, покрыта роговыми чешуями и ороговевшим эпителием и не участвует в дыхании. Акт дыхания у ящериц происходит путем расширения и сжатия грудной клетки под действием специальной мускулатуры.
Чтобы противостоять этому, следует помнить, что что-то вроде 99% нейронов в мозге и спинном мозге нельзя классифицировать как сенсорный или моторный, но можно рассматривать как «промежуточную сеть», идущую между строго сенсорным или строго двигательным нервом клетки. Подразумевается, что то, что начинается с отжига сенсорных клеточных аксонов и заканчивается как сокращение мышц, подвержено огромному раздуванию в нервной системе до того, как обращение этого процесса сходится, чтобы произвести поведение.
Между ощущением и действием мозг даже самого примитивного позвоночного имеет свою собственную жизнь. Разветвление сенсорной информации на несколько пунктов назначения хорошо иллюстрируется обзором визуальных путей, данных Эббессоном. Это показало, что начальная проекция волокон из сетчатки глаза проходит до пяти или шести разных точек во всех классах позвоночных. Оставляя в стороне большинство из них, которые могут иметь довольно узкие полоски функции, есть еще два основных визуальных пути. Один начинается с проекции от глаза к зрительному тектому в среднем мозге и обычно считается доминирующим в представлении нижних позвоночных, а другой начинается с проекции сетчатки на таламус.
Особенности дыхательной системы птиц. Механизм дыхания у птиц
У птиц воздух через ноздри и хоаны попадает в ротовую полость. Далее дыхательный путь идет через гортанную щель и верхнюю гортань и далее в трахею. В связи с удлинением шеи у птиц трахея намного длиннее, чем у пресмыкающихся. Войдя в полость тела, она на уровне сердца разделяется на два бронха, впадающих в правое и левое легкие и там сильно ветвящихся. Нижняя часть трахеи и начальные участки бронхов образуют характерную только для птиц нижнюю гортань- голосовой аппарат. Парные легкие птиц небольшие по размерам и плотно прикреплены к ребрам по бокам позвоночного столба. Легкие имеют плотное губчатое строение.
Все позвоночные используют это двойное видение, возможно, потому что один путь обнаруживает движущиеся объекты, а другой анализирует неподвижные узоры. Но в каждом случае участвуют несколько ядер в таламусе, и чрезвычайно сложные трансформации и абстракции могут иметь место в полушариях головного мозга даже у нижних позвоночных, таких как акулы.
Использование по меньшей мере двух параллельных, но взаимодополняющих интерпретаций сенсорного ввода для одной модальности может быть правилом, а не. Акустическая информация от каждого уха млекопитающих проходит до двух ядер в стволе мозга, а из них идет либо прямо к таламусу, либо по отдельному маршруту - в среднем мозгу, а затем проходит через таламус в слуховую кору. Аналогично хорошо известно, что у млекопитающих имеется по крайней мере два проекции поверхности тела на коре. Движение автотранспорта также проходит по двойным или тройным переулкам, прежде чем сходится к мышцам, основное различие между пирамидальной дорожкой, идущей от моторной коры к спинному мозгу, и экстрапирамидальной цепью через не кортикальные структуры переднего мозга, а также срединный мозг и мозговой мозг, с обходами, взятыми обоими, чтобы забрать разработки назначения в мозжечке.
От легких у птиц отходят воздушные мешки. Акт дыхания птиц осуществляется движениями грудной клетки. Благодаря сокращениям реберных мышц и подвижности ребер тело грудины отходит от позвоночного столба, объем полости тела увеличивается, растягиваются эластичные воздушные мешки и происходит засасывание воздуха в легкие и в задние воздушные мешки (задние грудные и брюшные); в растягивающиеся передние воздушные мешки (шейные, межключичный, передние грудные) при этом засасывается воздух из легких. При выдохе грудина придвигается к позвоночному столбу, тело грудины и внутренние органы сдавливают воздушные мешки, и уже прошедший через легкие воздух из передних воздушных мешков выдавливается в трахею, выводится наружу, а свежий воздух из задних воздушных мешков нагнетается в легкие. Так у птиц практически непрерывно - и во время вдоха и во время выдоха - через легкие в одном направлении идет поток свежего воздуха.
Понятно, что большая часть богатства и разнообразия центральной нервной системы теряется, если мы конденсируем функцию до ощущения, действия и корреляции между ними: нужно использовать термины типа «сенсорный» и «мотор» не как метки для элементарных категорий, а как синонимы проблем, для практического решения которых требуется эволюция сложного органа, как мозг, и чья интерпретация в нейропсихологической теории по-прежнему носит предварительный характер.
Гипотезы филогенетических изменений в организации мозга. Принимая во внимание ограничения основных понятий, мы теперь можем более подробно рассказать о том, как организация мозга позвоночных может меняться от класса к классу. Наиболее распространенная форма доктрины энцефализации предполагает, что функции управления поведением постепенно концентрируются на более высоких уровнях мозга. Это понятие было применено к эволюции позвоночных от беспозвоночных, эволюция.
Такое своеобразное строение дыхательной системы птиц обеспечивает интенсивное насыщение крови кислородом. У млекопитающих воздух поступает через наружные ноздри в обонятельную полость, а оттуда через хоаны в глотку и гортань. В гортани располагаются голосовые связки. Гортань переходит в трахею- длинную трубку, состоящую из незамкнутых на дорзальной стороне хрящевых колец. В грудной клетке трахея разделяется на два бронха, направляющихся в легкие. В легких бронхи многократно ветвятся на все меньшие по диаметру трубочки; самые мелкие из них оканчиваются тонкостенными пузырьками - альвеолами. В стенках альвеол располагаются кровеносные капилляры; именно здесь и происходит газообмен. Альвеолярная структура легких свойственна только млекопитающим.
Последовательные классы позвоночных, а также различия между заказами млекопитающих и внутри них. С млекопитающими энцефализацией считается кортикализация. Часто считается, что приматы, будучи более продвинутыми, более широко используют свою кору, чем грызуны или плотоядные, а также, что человек более зависим от кортикальных функций, чем просимий, обезьян или обезьян, в этой последовательности других приматов.
Возникновение гипотезы о гипотезе состоит в том, что дело любой части центральной нервной системы можно рассматривать в широком смысле как интерпретацию сенсорной информации, а также контроль и организацию поведения. Как только это будет сделано, есть тенденция хотеть назначить какое-то подразделение мозга как «ответственное» всех остальных, и это создает предпосылки для филогенетических взятий исполнительной власти. Преобладание определенного уровня мозга часто связано с обязанностями корреляции, интеграции или ассоциации, поэтому мы можем найти ссылки на средний мозг в качестве доминирующего центра корреляции или на таламус как наиболее развитый механизм ассоциации в определенном группа позвоночных. «Миграция доминирующего контрольного центра», по-видимому, связана с зачисткой коррелятивных и ассоциативных активов рыбы и среднего земного амфибия рептильным и высшим позвоночным передним мозгом.
Легкие свободно висят на бронхах в грудной полости. Каждое легкое разделяется на доли, количество которых варьирует у разных видов млекопитающих. Грудная полость млекопитающих четко отделена от брюшной полости сплошной мускулистой перегородкой - диафрагмой. Акт дыхания осуществляется синхронными движениями грудной клетки и диафрагмы. При вдохе объем грудной полости резко возрастает благодаря расширению грудной клетки и уплощению диафрагмы; легкие при этом расширяются, насасывая воздух. При выдохе стенки грудной клетки сближаются, а диафрагма куполом вдается в грудную полость. При этом общий объем грудной полости уменьшается, давление в ней возрастает и легкие сжимаются, воздух из них выталкивается.
Особенности процесса дыхания у человека и высших животных
Источником энергии в организме служат питательные вещества. Основной биохимической реакцией, освобождающей энергию этих веществ, является окисление, сопровождающееся расходованием кислорода и образованием углекислого газа. В организме человека нет запасов кислорода, поэтому его непрерывное поступление жизненно необходимо. Прекращение доступа кислорода в клетки организма приводит к их гибели. Образующийся при окислении веществ углекислый газ должен быть удален из организма, так как его накопление в значительном количестве опасно для жизни. Обмен кислорода и углекислого газа между организмом и окружающей средой называется дыханием. У человека и высших животных процесс дыхания осуществляется в следующей последовательности: обмен воздуха между атмосферой и альвеолами легких, обмен газов между альвеолами легких и кровью (внешнее дыхание), транспорт газов кровью, обмен газов между кровью и тканями (внутреннее, тканевое дыхание). Органы дыхания включают воздухоносные пути и легкие. Нос, носоглотка, гортань, трахея, бронхи и бронхиолы служат для проведения воздуха в альвеолы легких, где осуществляется газообмен.
Полость носа
Дыхательная система начинается полостью носа, которая образована костями лицевой части черепа и хрящами. Входом в полость носа служат ноздри, а выходом хоаны – отверстия, сообщающие его полость с носоглоткой. Стенки носовой полости образуют неровный рельеф, за счет чего увеличивается площадь контакта воздуха с наружным слоем клеток слизистой оболочки. Многие из них имеют реснички. Вдыхаемый воздух, проходя через носовую полость, согревается кровью, протекающей по многочисленным, пронизывающим оболочку сосудам, и, кроме того, соприкасаясь со слизистой, увлажняется и частично очищается от пыли, микробов и ряда других примесей. Из носовой полости воздух попадает в носоглотку, далее в ротовую часть глотки и затем в гортань.
Гортань
Гортань имеет сложное строение, так как служит не только для проведения воздуха, но и для образования звуков. Гортань состоит из хрящей различной формы, соединенных связками и суставами, приводимыми в движение мускулатурой. Скелет гортани образован непарными (щитовидный, перстневидный и надгортанный) и парными (черпаловидные, рожковидные и клиновидные) хрящами. Самый крупный – щитовидный хрящ – находится спереди. Полость гортани выстлана слизистой оболочкой, образующей голосовые связки. Между свободными краями связок расположена в продольном направлении голосовая щель.
Напряжение и расслабление голосовых связок регулируется специальными мышцами. В спокойном состоянии, когда человек молчит, голосовая щель открыта и имеет вид равнобедренного треугольника. Во время разговора или пения голосовые связки натягиваются, сближаются и при прохождении выдыхаемого воздуха вибрируют, производя звук. Однако окончательное формирование звука происходит в полостях рта, носа, глотки и зависит от положения языка, губ, нижней челюсти. Высота звука определяется длиной голосовых связок: чем длиннее связки, тем меньше частота их колебания и тем ниже голос. Вход в гортань прикрывает надгортанник, препятствующий попаданию пищи в дыхательные пути. Книзу гортань переходит в трахею (дыхательное горло).
Трахея
Трахея у взрослого человека имеет форму трубки длиной 10-13 см и служит для прохождения воздуха в легкие и обратно. Она образована 16-20 полукольцами из гиалинового хряща, которые придают жесткость и не дают спадаться трахее. Между собой хрящевые полукольца соединены плотной соединительной тканью. Сзади между концами полуколец находится соединительнотканная перепонка. Благодаря наличию эластичных волокон в соединительной ткани между полукольцами, трахея может удлиняться при движении гортани вверх и укорачивается при ее опускании. Полость трахеи выстлана мерцательным эпителием, реснички которого перемещают попадающие с воздухом частицы пыли вместе со слизью вверх в глотку, где они проглатываются. Нижний конец трахеи делится на две более тонкие трубки – бронхи (правый и левый). Место деления называется бифуркацией трахеи.
Бронхи
Бронхи постепенно разветвляются на более мелкие, доходя до тонких и тончайших веточек – бронхиол, диаметр которых не превышает долей миллиметра. В целом разветвление бронхов образует густую сеть – бронхиальное дерево. Крупные бронхи, как и трахея, состоят из хрящевых колец, связанных между собой соединительной тканью. В бронхиолах хрящевой скелет отсутствует, однако из стенки не спадается, так как состоят из гладкомышечных волокон. Бронхиолы – последние элементы воздухоносных путей.
Легкие
Легкие представляют собой парные губчатые органы конусовидной формы. Ткань легкого образована бронхиолами и множеством крошечных легочных пузырьков – альвеол, которые имеют вид полушаровидных выпячиваний бронхиол. Стенки альвеол состоят из одного слоя эпителиальных клеток, окруженных густой сетью кровеносных капилляров. Из нутрии альвеолы покрыты жидким поверхностно-активным веществом (сурфактантом), ослабляющим силы поверхностного натяжения и предупреждающим полное спадение альвеол во время выдоха. Суммарная толщина стенок альвеолы и капилляра составляет несколько микрометров. Благодаря такому строению кислород легко проникает из альвеолярного воздуха в кровь, а углекислый газ – из крови в альвеолы. Диаметр альвеол составляет в среднем 0,3 мм, однако благодаря тому, что в легких альвеол насчитывается до 300 млн. и их суммарная поверхность у взрослого человека равна 50-100 квадратных метров, газообмен в легких совершается чрезвычайно быстро. Легкие (правое и левое) расположено в грудной клетке и тесно прилегает к ее стенкам. Поверхность легких покрыта особой оболочкой – плеврой, состоящей из двух листков: наружный листок выстилает внутреннюю поверхность грудной клетки, а внутренний покрывает поверхность легкого. Между листками сохраняется герметически замкнутое щелевидное пространство, называемое плевральной полостью. В ней содержится небольшое количество жидкости, увлажняющей листки плевры и способствующей скольжению их относительно друг друга.
Основные преобразования в дыхательной системе хордовых животных.
Эволюция дыхательной системы.
Дыхательная система выполняет функцию газообмена между организмом и окружающей средой.
У животных с низким обменом веществ газовый обмен происходит путем диффузии через всю поверхность тела. Увеличение размеров тела, повышение интенсивности обменных процессов в организме привело к развитию специального дыхательного аппарата.
Эволюционные преобразования в дыхательной системе хордовых.
Усиление главной дыхательной функции:
1. увеличение поверхности газообмена;
2. дифференцировка воздухоносных и респираторных отделов;
3. совершенствование механизмов дыхания: появление грудной
клетки, дыхательной мускулатуры.
2. Расширение числа выполняемых функций: очищение, согревание, увлажнение воздуха; терморегуляция, звукообразование.
3. Субституция функций: дыхание с помощью жабр у наземных позвоночных замещается газообменом в легких.
4. Смена функций: плавательный пузырь древних кистеперых рыб преобразуется в орган дыхания.
5. Разделение функций и органов:
1. у наземных позвоночных отделение дыхательных путей от
первичной пищеварительной трубки;
2. в легких позвоночных разделение воздухоносных и респираторных отделов.
В процессе эволюции устанавливается морфофунациональная взаимосвязь дыхательной и кровеносной систем (динамические координации), что выражается в расположении сосудов в органах дыхания, строении стенок альвеол и капилляров (аэро-гематический барьер), наличии транспортных систем для переноса газов, присутствии дыхательных пигментов в крови.
Функции нервной системы.
Главной функцией нервной системы является внедрение воздействия на организм извне, сопровождающейся приспособительной реакцией человеческого организма. Головной мозг состоит из ствола и переднего мозга. Каждый отдел мозга отвечает за выполнение определенных задач. Рассмотрим функции центральной нервной системы:
Так как передний мозг разделяется на конечный и промежуточный, следовательно, каждый несет в себе какие-то функции. Так, гипоталамус, таламус и лимбическая система входят в состав промежуточного. Первый является центром витальных потребностей (либидо, голод), эмоций. Таламус осуществляет первичную обработку информации, ее фильтрацию. Лимбическая система отвечает за эмоционально-импульсивное поведение личности.
В состав данной нервной системы входят клетки, называемые нейроглии. Они выполняют опорную функцию, принимают участие в метаболизме клеток нервной системы.
В спинном мозге имеется белое вещество, что образует проводящие пути. Они соединяют спинной и главный мозг, отдельные сегменты данного мозга друг с другом. Пути выполняют проводниковую, рефлекторную функцию.
Анализаторы играют роль отражателей в сознании человека внешнего материального мира.
Деятельность коры головного мозга являет собой высшую нервную деятельность и выполняет условно - рефлекторную функцию.
Основные функции центральной нервной системы заключаются в осуществлении простых и сложных отражательных реакций, называемых рефлексами.
ЦНС с конечностями и органами соединяет периферическая нервная система. Она не защищена костями, что говорит о том, что она может быть подвергнута действиям токсинов и механических повреждений.
Функции периферической нервной системы
ПНС разделяется на вегетативную и соматическую, каждая из которых выполняет определенные функции. Соматическая нервная система ответственна за координацию движений и за получение стимулов, приходящих из внешнего мира. Она регулирует деятельность, что контролирует сознание человека.
Вегетативная, в свою очередь, выполняет защитную функцию в случае, если надвигается опасность или стрессовая ситуация. Отвечает за кровяное давление и пульс. Когда же человек волнуется, она, зафиксировав чувство волнения, повышает уровень адреналина.
Парасимпатическая система, входящая в состав вегетативной, выполняет свои функции, когда индивид находится в состоянии покоя. Она ответственна за сужение зрачков, стимуляцию мочеполовой и пищеварительной системы.
И все-таки, какие функции выполняет нервная система?
Получение информации об окружающем мире человека и состоянии организма.
Передача этой информации в головной мозг.
Координация сознательногофункции центральной нервной системы телодвижения.
Координация и регулирование сердечного ритма, температуры и пр.
Особенности строения и функций различных отделов головного мозга у рыб, амфибий, рептилий, птиц, млекопитающих и человека.
Различия ихтиопсидного, зауропсидного и маммилийного типов мозгов.
Основные направления эволюции нервной системы у различных представителей позвоночных.
Гомология основных отделов головного мозга у различных представителей позвоночных.
Терминальный тератогенный фактор.
Онтофилогенетически обусловленные пороки развития дыхательной системы у человека. Примеры.
Основные врожденные филогенетически обусловленные пороки развития головного и спинного мозга у человека. Примеры.
В связи с выходом на сушу и более активной жизнедеятельностью характерной для высших позвоночных, все отделы мозга пресмыкающихся достигают более прогрессивного развития.
1. Передний мозг значительно преобладает над другими отделами. Мантия остается тонкой, но на ее поверхности местами появляются медиальное и латеральное скопления нервных клеток – серое вещество, представляющее зачаточную кору больших полушарий. У рептилии кора еще не играет роли высшего отдела мозга, она является высшим обонятельным центром. Но в процессе филогенеза, разрастаясь и принимая другие виды чувствительности, помимо обонятельной, она привела к возникновению коры головного мозга млекопитающих. Полушария переднего мозга пресмыкающихся полностью прикрывают промежуточный мозг. Роль высшего интегративного центра выполняют полосатые тела (зауропсидный тип мозга)
2. Промежуточный мозг образован зрительными буграми и подбугровой областью. На дорсальной его стороне находится эпифиз и особый теменной орган, имеющий глазоподобное строение у ящериц. На вентральной стороне находится гипофиз.
3. Средний мозг довольно большой, имеет вид двухолмия. Это центр зрительных восприятий, приобретающий большое значение для наземных животных.
4. Мозжечок имеет вид полукруглой пластинки, развит слабо, но лучше, чем у амфибий, в связи с усложнением координации движений.
5. Продолговатый мозг образует резкий изгиб, характерный для высших позвоночных. От его ядер берут начало черепно-мозговых нервов.
Всего у рептилий 12 пар черепно-мозговых нервов.
ГОЛОВНОЙ МОЗГ ПТИЦ

Эволюция головного мозга позвоночных: а - рыба; б - земноводное; в - пресмыкающееся; г - млекопитающее; 1 - обонятельные доли; 2 - передний мозг; 3 - средний мозг; 4 - мозжечок; 5 - продолговатый мозг; 6 - промежуточный мозг
1.Передний мозг развит хорошо, полушария имеют значительную величину, частично прикрывают промежуточный мозг. Но увеличение полушарий происходит за счет развития полосатых тел (зауропсидный тип мозга), а не коры. Обонятельные доли очень малы, так как обоняние теряет ведущее значение.
2 Промежуточный мозг мал, прикрыт полушариями переднего мозга. На дорсальной стороне его находится эпифиз (развит слабо), а на вентральной – гипофиз.
3. Средний мозг довольно большой, за счет крупных зрительных долей (двухолмие), что связано с прогрессивным развитием зрения.
4. Мозжечок сильно развит в связи со сложной координацией движений при полете. Он имеет поперечную исчерченность, и свою кору.
5. Продолговатый мозг содержит скопление нервных клеток в виде ядер, от которых берут начало черепно-мозговые нервы от 5-й до 12-й пары.
Всего 12 пар черепно-мозговых нервов.
ГОЛОВНОЙ МОЗГ МЛЕКОПИТАЮЩИХ
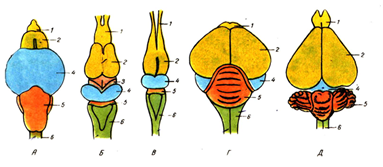
Эволюция головного мозга позвоночных: а - рыба; б - земноводное; в - пресмыкающееся; г - млекопитающее; 1 - обонятельные доли; 2 - передний мозг; 3 - средний мозг; 4 - мозжечок; 5 - продолговатый мозг; 6 - промежуточный мозг
1 Передний мозг достигает особенно больших размеров, прикрывая остальные отделы мозга. Его увеличение происходит за счет коры, которая становится главным центром высшей нервной деятельности (маммальный тип мозга). Площадь коры увеличивается за счет образования извилин и борозд. Спереди от больших полушарий у большинства млекопитающих (кроме китообразных, приматов и, в том числе, человека) расположены крупные обонятельные доли, что связано с большим значением обоняния в жизни зверей.
2 Промежуточный мозг, образованный зрительными буграми (thalamus) и подбугровой областью (hypothalamus), скрыт полушариями переднего мозга. На дорсальной его стороне находится эпифиз, а на вентральной – гипофиз.
3 Средний мозг прикрыт полушариями переднего мозга, отличается сравнительно небольшими размерами и представлен не двухолмием, а четверохолмием. Полость среднего мозга, или сильвиев водопровод, представляет собой лишь узкую щель.
4 Мозжечок сильно развит и имеет более сложное строение; состоит из центральной части – червя с поперечными бороздами и парных полушарии. Развитие мозжечка обеспечивает сложные формы координации движений.
5 Продолговатый мозг частично прикрыт мозжечком. Отличается от представителей других классов тем, что потоком четвертого желудочка обособляются продольные пучки нервных волокон – задние ножки мозжечка, а на нижней поверхности имеются продольные валики – пирамиды. От головного мозга отходит 12 пар черепно-мозговых нервов
48. 50. Филогенетически сложившиеся типы и формы иммунного ответа. Характеристика особенностей иммунной системы позвоночных.
Филогенез иммунной системы.
Иммунная система осуществляет защиту организма от проникновения в организм генетически чужеродных тел: микроорганизмов, вирусов, чужих клеток, инородных тел. Ее действие основано на способности отличать собственные структуры от генетически чужеродных, уничтожая их.
В эволюции сформировалось три главных формы иммунного ответа:
1) 1.Фагоцитоз, или неспецифическое уничтожение чужеродного материала;
2) 2.Клеточный иммунитет, основанный на специфическом распознавании и уничтожении такого материала Т-лимфоцитами;
3) 3.Гуморальный иммунитет, осуществляемый путем образования потомками В-лимфоцитов, так называемыми, плазматическими клетками иммуноглобулинов и связывания ими чужеродных антигенов.
В эволюции выделяют три этапа формирования иммунного ответа:
1. 1. Квазииммунное (лат наподобие) распознавание организмов своих и чужеродных клеток. Этот тип реакции наблюдается от кишечнополостных до млекопитающих. Эта реакция не связана с выработкой иммунных тел, и при этом не формируется иммунной памяти, то есть еще не происходит усиления иммунной реакции на повторное проникновение чужеродного материала.
2. 2. Примитивный клеточный иммунитет обнаружен у кольчатых червей и иглокожих. Он обеспечивается целомоцитами – клетками вторичной полости тела, способными уничтожать чужеродный материал. На этом этапе появляется иммунологическая память.
3. 3. Система интегрального клеточного и гуморального иммунитета . Для нее характерны специфические клеточные и гуморальные реакции на чужеродные тела, наличие лимфоидных органов иммунитета, образование антител. Такого типа иммунная система не характерна для беспозвоночных.
Круглоротые способны формировать антитела, но вопрос о наличии у них вилочковой железы, как центрального органа иммуногенеза, является пока открытым. Впервые тимус обнаруживается у рыб.
Эволюционные предшественники лимфоидных органов млекопитающих – тимус, селезенка, скопление лимфоидной ткани обнаруживаются в полном объеме у амфибий. У низших позвоночных (рыбы, амфибии) вилочковая железа активно выделяет антитела, что характерно для птиц и млекопитающих.
Особенность иммунного ответа птиц состоит в налиции особоги лимфоидного органа – фабрициевой сумки. В этом органе образуются В-лимфоциты, которые после антигенной стимуляции способны трансформироваться в плазматические клетки и вырабатывать антитела.
У млекопитающих органы иммунной системы разделяют на два типа: центральные и периферические. В центральных органах созревание лимфоцитов происходит без существенного влияния антигенов. Развитие периферических органов, наоборот, непосредственно зависит от антигенного воздействия – лишь при контакте с антигеном в них начинаются процессы размножения и дифференциации лимфоцитов.
Центральными органами иммуногенеза у млекопитающих являются тимус, где происходит образование и размножение Т-лимфоцитов, а также красный костный мозг, где образуются и размножаются В-лимфоциты.
На ранних стадиях эмбриогенеза и желточного мешка в тимус и красный костный мозг мигрируют стволовые лимфотические клетки. После рождения источником стволовых клеток становится красный костный мозг.
Периферическими лимфоидными органами являются: лимфоузлы, селезенка, миндалины, лимфоидные фолликулы кишечника. К моменту рождения они еще практически не сформированы и образование в них лимфоцитов начинается только после антигенной стимуляции, после того, как они заселяются Т- и В-лимфоцитами из центральгых органов иммуногенеза.
49. 51. Онтогенез, его типы и периодизация.
Онтогенез, или индивидуальное развитие, – это совокупность преобразований, происходящих в организме от момента образования зиготы до смерти. Термин «онтогенез» впервые введен биологом Э.Геккелем в 1866 г. (от греч. онтос- существо и генезис- развитие).
Учение об онтогенезе – это один из разделов биологии, который изучает механизмы, регуляцию и особенности индивидуального развития организмов.
Знание онтогенеза имеет не только общетеоретическое значение. Оно необходимо врачам для понимания особенностей течения патологических процессов в разные возрастные периоды, профилактики заболеваний, а также для решения социально- гигиенических проблем, связанных с организацией труда и отдыха людей различных возрастных групп.
Различают 2 типа онтогенеза: непрямой и прямой. Непрямой протекает в личиночной форме. Личинки ведут активный образ жизни, сами себе добывают пропитание. Для осуществления жизненных функций у личинок имеется ряд провизорных (временных) органов, отсутствующих у взрослых организмов. Этот тип развития сопровождается метаморфозом (превращением) -анатомо-физиологической перестройкой организма. Он свойствен различным группам беспозвоночных (губкам, кишечнополостным, червям, насекомым) и нисшим позвоночным (амфибиям).
Прямое развитие может протекать в неличиночной форме или быть внутриутробным. Неличиночный тип развития имеет место у рыб, пресмыкающихся, птиц, а также беспозвоночных, яйцеклетки которых богаты желтком - питательным материалом, достаточным для завершения онтогенеза. Для питания, дыхания и выделения у зародышей также развиваются провизорные органы.
Внутриутробный тип развития характерен для млекопитающих и человека. Их яйцеклетки почти не содержат питательного материала, и все жизненные функции осуществляются через материнский организм. В связи с этим у зародышей имеются провизорные органы – зародышевые оболочки и плацента, обеспечивающая связь организма матери и плода. Это наиболее поздний в филогенезе тип онтогенеза, и он обеспечивает наилучшим образом выживание зародышей.
Онтогенез включает в себя ряд преемственно связанных и в основных чертах генетически запрограммированных периодов:
1. Предэмбриональный (он же проэмбриональный, или предзиготный период, или прогенез);
2. Эмбриональный (или антенатальный для человека) период;
3. Постэмбриональный (или постнатальный для человека) период.
a. 52. Общая характеристика предзиготного периода, стадии эмбрионального развития. Критические периоды. Тератогенные факторы.
ПРЕДЗИГОТНЫЙ ПЕРИОД
Этот период протекает в организме родителей и выражается в гаметогенезе – образовании зрелых яйцеклеток и сперматозоидов.
В настоящее время известно, что в этот период происходит ряд процессов, имеющих прямое отношение к ранним стадиям эмбрионального развития. Так, в ходе созревания яйцеклеток в пахинеме мейоза наблюдается амплификация генов (образование многочисленных копий), отвечающих за синтез р-РНК, с последующим выделением их из ДНК и накоплением вокруг ядрышек. Эти гены включаются в транскрипцию на ранних стадиях эмбриогенеза, обеспечивая накопление р-РНК, участвующей в образовании рибосом. Кроме того, в предзиготном периоде происходит также накопление как бы впрок и-РНК, включающейся в биосинтез белка только на ранних стадиях дробления зиготы.
Во время овогенеза в яйцеклетках идет накопление желтка, гликогена и жиров, которые расходуются в процессе эмбриогенеза.
По количеству содержания желтка (lecithos) яйцеклетки могут быть:
· олиголецитальными (маложелтковыми);
· мезолецитальными (со средним количеством желтка);
· полилецитальными (многожелтковые).
По характеру распределения желтка в цитоплазме яйцеклетки бывают:
· изолецитальные (греч. Isos – равный, желток распределен в клетке равномерно);
· телолецитальные (греч. thelos – конец, желток смещен ближе к вегетативному полюсу, а клеточное ядро – к анимальному);
· центролецитальные (желток располагается в центральной части яйцеклетки)
Изолецитальные клетки характерны для ланцетника и млекопитающих, телолецитальные – для амфибий (умеренно телолецитальные, для рептилий и птиц – резкотелолецитальные), центролецитальные – для насекомых.
Яйцеклетки некоторых видов животных еще до оплодотворения приобретают билатеральную симметрию, однако она еще неустойчива и может в дальнейшем переориентироваться.
У многих видов животных еще до оплодотворения начинается сегрегация (перераспределение) органоидов и включений в яйцеклетках; отмечается скопление гликогена и и РНК на анимальном полюсе, комплекса Гольджи и аскорбиновой кислоты - на экваторе. Сегрегация продолжается и после оплодотворения.
ЭМБРИОНАЛЬНЫЙ ПЕРИОД
Эмбриональный период начинается с зиготы и заканчивается либо выходом молодых особей из яйцевых оболочек, либо рождением нового организма. Этот период состоит из стадий: зиготы, дробления, гаструляции и гисто- и органогенеза.
ХАРАКТЕРИСТИКА СТАДИЙ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
НА ПРИМЕРЕ ЧЕЛОВЕКА.
После оплодотворения наступает первая стадия эмбрионального развития - стадия зиготы (стадия одноклеточного зародыша). Зигота, будучи одной клеткой, имеет потенции к развитию целостного многоклеточного организма, т.е. обладает тотипотентностью.
Стадия дробления: начиная с этой стадии, зародыш становится многоклеточным, но по размерам практически не превышает зиготу. Дробление заключается в том, что хотя клетки делятся митозом, они не вырастают до размеров материнских клеток, т.к. у них отсутствует гетеросинтетическая интерфаза, а период G1 аутосинтетической интерфазы приходится на телофазу предшествующего деления. Стадия дробления заканчивается образованием бластулы. Первые бластомеры, как и зигота, обладают свойством тотипотентности, что служит основой рождения монозиготных (однояйцевых) близнецов.
У человека бластула образуется на 6-7 день развития и имеет вид пузырька (бластоциста), стенки которого образованы одним слоем клеток - трофобластом, выполняющих функции питания и выделения. Внутри пузырька имеется скопление клеток - эмбриобласт, из которого в дальнейшем развивается тело зародыша.
На стадия гаструляции (у человека с 7 по 19 день) происходит образование зародышевых листков (эктодермы, энтодермы и мезодермы), и закладывается комплекс осевых органов (хорда, нервная трубка и кишечная трубка).
В период гистогенеза и органогенеза идет закладка временных (провизорных) и окончательных (дефинитивных) органов. У позвоночных животных, в том числе и у человека, провизорные органы называются зародышевыми оболочками. Для всех позвоночных характерно развитие желточного мешка. У рыб, амфибий, рептилий и птиц он содержит желток и выполняет трофическую и кроветворную функции. У истинно наземных животных, кроме желточного мешка, имеется также амнион, наполненный жидкостью, создающей водную среду для развития зародыша. Позвоночные, имеющие амнион (рептилии, птицы и млекопитающие), называются амниотами , а не имеющие его - анамниями (рыбы, амфибии).
У рептилий и птиц, кроме желточного мешка и амниона, закладываются: аллантоис (мочевой мешок, накапливающий мочевину) и серозная оболочка (обеспечивает дыхание зародыша). У млекопитающих вместо серозной оболочки образуется хорион (ворсинчатая оболочка), который обеспечивает зародышу питание, дыхание и выделение. Хорион образуется из трофобласта и соединительной ткани. Со стадии плацентации он участвует в образовании плаценты. Амнион содержит околоплодные воды. В желточном мешке образуются первые кровеносные сосуды и первые клетки крови. Аллантоис у млекопитающих и человека определяет место расположения плаценты.
Гисто- и органогенез у человека начинается на четвертой неделе и заканчивается к рождению.
Вначале из так называемой первичной эктодермы вычленяются клетки, образуя нервную пластинку, из которой в дальнейшем развиваются все органы нервной системы и часть органов чувств. Из оставшейся вторичной эктодермы закладываются эпидермис и его производные - сальные, потовые, молочные железы, ногти, волосы и некоторые другие образования.
Из энтодермы формируются: эпителий желудочно-кишечного тракта, дыхательных путей, печень и поджелудочная железа.
Из мезодермы - скелет, поперечнополосатая и гладкая мускулатура, сердечно-сосудистая система и основная часть мочеполовой системы.
КРИТИЧЕСКИЕ ПЕРИОДЫ РАЗВИТИЯ
В 1921 г. Стоккард Ц.Р. положил начало представлениям о так называемых критических периодах развития животных организмов. Этой проблемой позже у нас в стране занимался Светлов П.Г., который в 1960 г. сформулировал теорию критических периодов развития, проверил ее экспериментально. Сущность ее состоит в том, что каждый этап развития зародыша начинается коротким периодом качественно новой перестройки, сопровождающемся детерминацией, пролиферацией и дифференцировкой клеток. В этот период наблюдается особая восприимчивость к различным повреждающим факторам среды – физическим, химическим и в ряде случаев – биологическим, которые могут ускорять, замедлять и даже приостанавливать развитие.
В онтогенезе человека выделяют следующие критические периоды: 1) гаметогенез; 2)оплодотворение; 3)имплантацию; 4)развитие комплекса осевых органов и формирование плаценты (3-8-я недели) 5) периоды дифференцировки того или иного органа или системы органов, (20-24-я недели);. 6)рождение; 7) период новорожденности (до 1 года); 8) половое созревание.


b. 53. Основные механизмы эмбриогенеза.
ОБЩИЕ МЕХАНИЗМЫ ЭМБРИОГЕНЕЗА
1.Деление клеток
2. Клеточная дифференцировка
3. Дифференциальная активность генов
4. Эмбриональная индукция
5. Межклеточные взаимодействия
6 Миграция клеток.
7.Гибель клеток
8. Клональный принцип развития
9. Рост.
10. Морфогенез.
В основе эмбрионального развития лежат разнообразные процессы /механизмы/, к которым относятся: клеточные деления, дифференцировка, эмбриональная индукция, межклеточные взаимодействия, миграция клеток, гибель клеток, клональный принцип развития, рост, морфогенез и дифференциальная активность генов.
1.Деление клеток лежит в основе пролиферации /разрастания клеток/ и является основным механизмом обеспечения роста, то есть увеличения массы и размеров тела. Кроме того, в ходе клеточных делений в ряде случаев происходит переключение генетических программ и, как следствие этого, - специализация клеток для выполнения определенных функций.
2. Клеточная дифференцировка - это процесс, когда из внешне однообразных клеток и их комплексов возникают специализированные клетки, отличающиеся от материнских морфологическими и функциональными особенностями. Этот процесс носит дивергентный /разнонаправленный/ характер. С биохимической точки зрения, дифференцировка - это выбор из некоего множества возможных путей биосинтеза какого-либо одного (например, клетками-предшественниками эритроцитов выбор пути синтеза гемоглобина, а клетками хрусталика глаза - белка кристалина). С морфологической точки зрения, дифференцировка выражается в приобретении специфических черт строения.
В результате дифференцировки развивается популяция высокоспециализированных клеток либо утративших ядра /эритроциты, ороговевшие клетки эпидермиса/, либо в клетках начинается синтез высокоспецифичных веществ, например, сократимых белков актина и миозина – в мышечных волокнах, определенных гормонов – в клетках желез внутренней секреции, и т. д.
Путь, по которому должна идти дифференцировка тех или иных клеток, генетически детерминирован /предопределен/. На стадии дробления детерминация клеток носит еще неустойчивый характер /лабильна/, и направление дифференцировки можно изменить. Это подтвердил в первой четверти ХХ века Ганс Шпеман в экспериментах на тритонах. Он пересаживал эктодермальные клетки, взятые у тритона одного вида, в энтодерму другого. И хотя клетки донора отличались по цвету от клеток реципиента, они развивались в те же зачатки, что и окружавшие их клетки реципиента. Если донорами были организмы, закончившие процесс гаструляции, то эктодермальные клетки нервной пластинки, пересаженные в кожу, давали зачаток нервной ткани, то есть путь их дифференцировки был уже предопределен.
Сейчас известны некоторые факторы, определяющие дифференцировку тканей. Самым ранним фактором, который проявляется уже на стадии бластулы, является сегрегация /лат. «отделение»/ цитоплазматических структур зиготы, в силу чего во время дробления в первые бластомеры попадают отличающиеся друг от друга участки цитоплазмы. Таким образом, видимо, незначительные качественные различия, имеющиеся в разных участках цитоплазмы ооцитов, влияют на судьбу бластомеров. Есть также данные, что дифференцировка многих тканей зародыша может происходить лишь при наличии некоторого критического числа клеток.
Основным механизмом дифференцировки клеток является дифференциальная активность генов.
3. Эмбриональная индукция – это влияние одной ткани или зачатка органа зародыша /индуктора/ на закладку других зачатков органов. Так, например, у позвоночных закладка хордо-мезодермального комплекса индуцирует /побуждает к развитию/ закладку нервной трубки.
Другой формой индуцирующих воздействий являются межклеточные взаимодействия.
4. Межклеточные взаимодействия осуществляются посредством щелевых контактов, где плазматическая мембрана одних клеток вступает в тесный контакт с плазмалеммой других клеток. В области этих контактов между клетками может передаваться слабый электрический ток, ионы неорганических веществ или даже относительно крупные молекулы органических веществ.
5 Миграция клеток. В ходе эмбриогенеза происходит миграция как отдельных клеток, так и их комплексов, на различные расстояния. Отдельные клетки обычно мигрируют при помощи амебоидного движения, обследуя при этом непрерывно свое окружение.
6.Гибель клеток (апоптоз) является необходимым процессом многих стадий развития зародыша. Так, разделению пальцев ног и рук предшествует гибель клеток, расположенных в межпальцевых промежутках.
7. Клональный принцип развития. Экспериментальным путем показано, что многим клеткам раннего зародыша не суждено участвовать в дальнейшем развитии. Многие структуры зародыша строятся из клеток, которые развиваются в ходе деление только отдельного, небольшого числа клеток.
8. Рост. Под ростом понимается увеличение массы тела и его размеров. Рост носит неравномерный характер, разные ткани и разные части зародыша растут с разной скоростью.
9. Морфогенез. Это процесс пространственного становления внешней и внутренней конфигурации частей тела и органов зародыша. Общепринятой теории, объясняющей механизмы этого процесса, пока нет. Наиболее подходящей является концепция позиционной информации , предложенная Л. Вольпертом /1975/,согласно которой клетки способны воспринимать позиционную информацию, которая содержит указание о местоположении клеток относительно других клеток и тем самым определяется план, в соответствии с которым происходит развитие зародыша.